

A small viral potassium ion channel with an inherent inward rectification
- PMID: 31010373
- PMCID: PMC6527081
- DOI: 10.1080/19336950.2019.1605813
A small viral potassium ion channel with an inherent inward rectification
Abstract
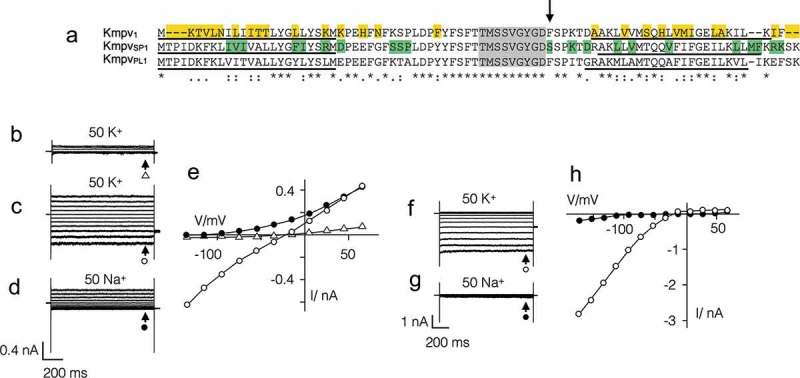
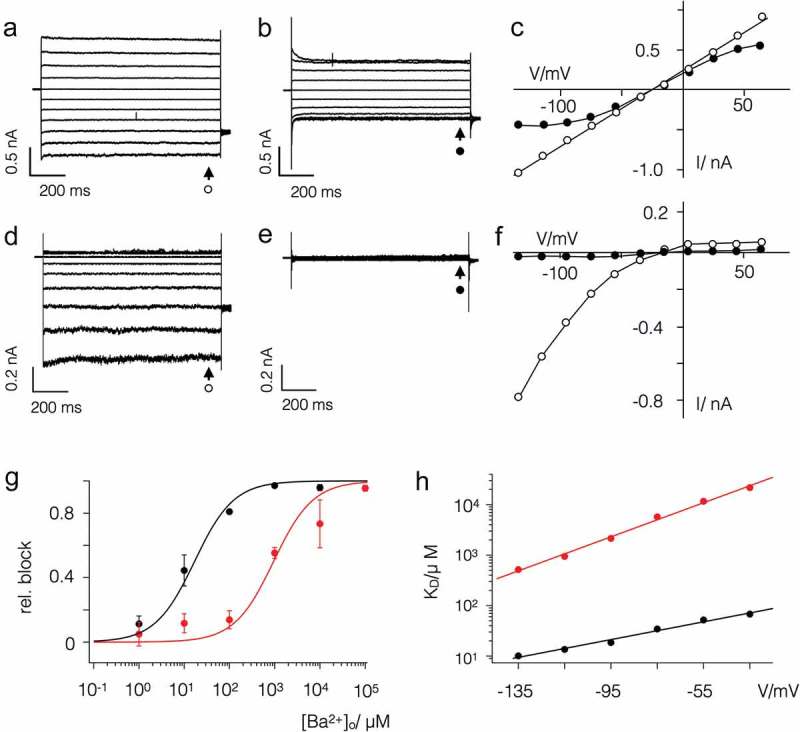
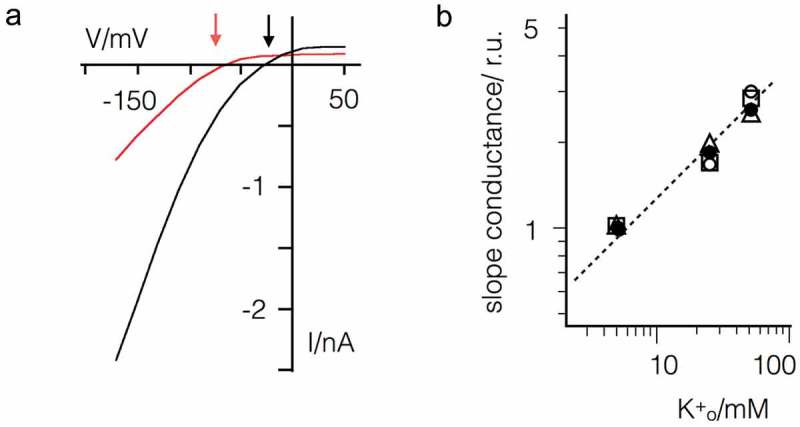
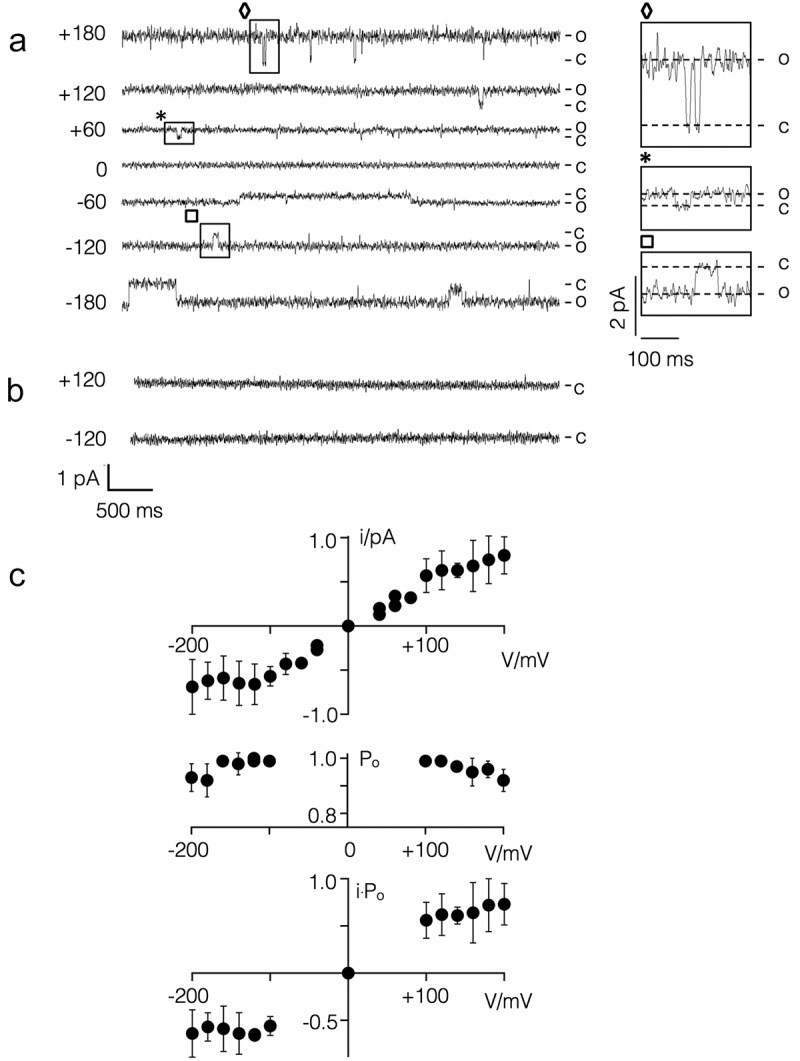
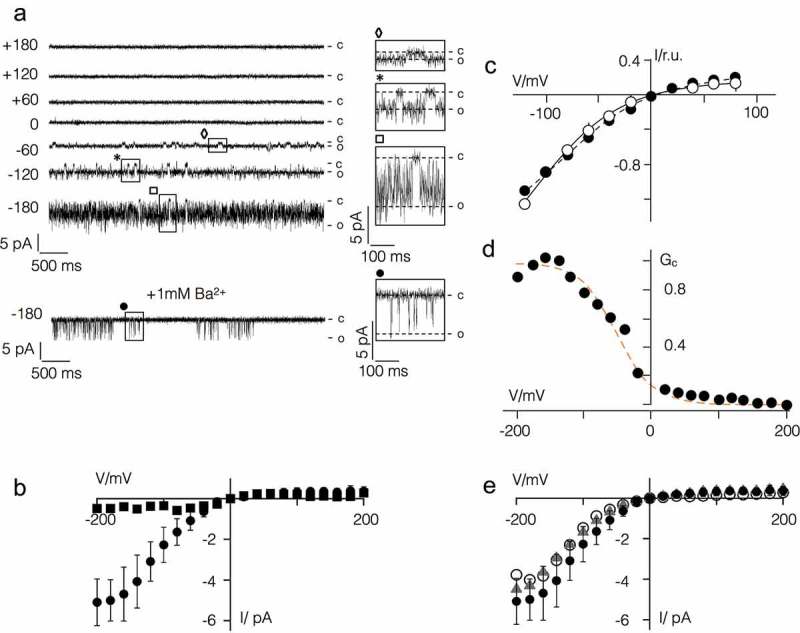
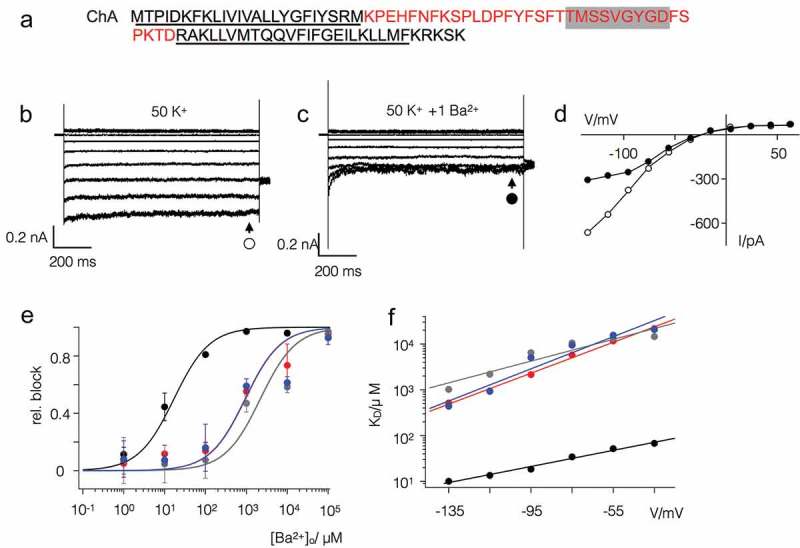
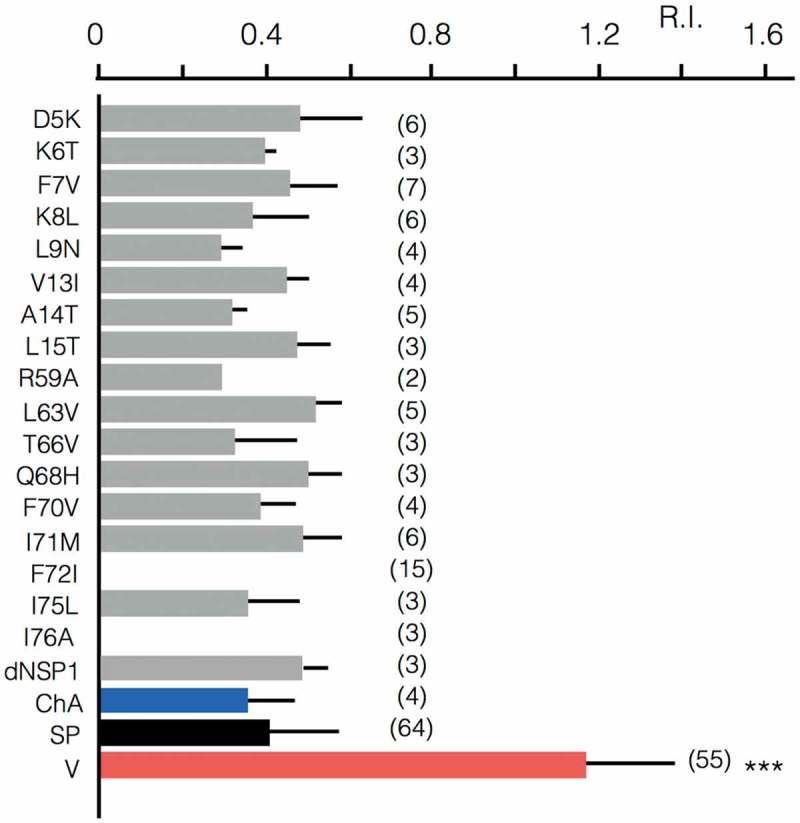
Some algal viruses have coding sequences for proteins with structural and functional characteristics of pore modules of complex K+ channels. Here we exploit the structural diversity among these channel orthologs to discover new basic principles of structure/function correlates in K+ channels. The analysis of three similar K+ channels with ≤ 86 amino acids (AA) shows that one channel (Kmpv1) generates an ohmic conductance in HEK293 cells while the other two (KmpvSP1, KmpvPL1) exhibit typical features of canonical Kir channels. Like Kir channels, the rectification of the viral channels is a function of the K+ driving force. Reconstitution of KmpvSP1 and KmpvPL1 in planar lipid bilayers showed rapid channel fluctuations only at voltages negative of the K+ reversal voltage. This rectification was maintained in KCl buffer with 1 mM EDTA, which excludes blocking cations as the source of rectification. This means that rectification of the viral channels must be an inherent property of the channel. The structural basis for rectification was investigated by a chimera between rectifying and non-rectifying channels as well as point mutations making the rectifier similar to the ohmic conducting channel. The results of these experiments exclude the pore with pore helix and selectivity filter as playing a role in rectification. The insensitivity of the rectifier to point mutations suggests that tertiary or quaternary structural interactions between the transmembrane domains are responsible for this type of gating.
Keywords: Ba2+ block; Kir channels; Viral K+ channel; inward rectification.
Figures
References
-
- Doyle DA, Cabral JM, Pfuetzner RA, et al. The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science. 1998;280:69–77. - PubMed
-
- Perozo E. New structural perspectives on K+ channel gating. Structure. 2002;10:1027–1029. - PubMed
-
- MacKinnon R. Potassium channels. FEBS Lett. 2003;555:62–65. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources