

Timing gone awry: distinct tumour suppressive and oncogenic roles of the circadian clock and crosstalk with hypoxia signalling in diverse malignancies
- PMID: 31014368
- PMCID: PMC6480786
- DOI: 10.1186/s12967-019-1880-9
Timing gone awry: distinct tumour suppressive and oncogenic roles of the circadian clock and crosstalk with hypoxia signalling in diverse malignancies
Abstract
Background: The circadian clock governs a large variety of fundamentally important physiological processes in all three domains of life. Consequently, asynchrony in timekeeping mechanisms could give rise to cellular dysfunction underpinning many disease pathologies including human neoplasms. Yet, detailed pan-cancer evidence supporting this notion has been limited.
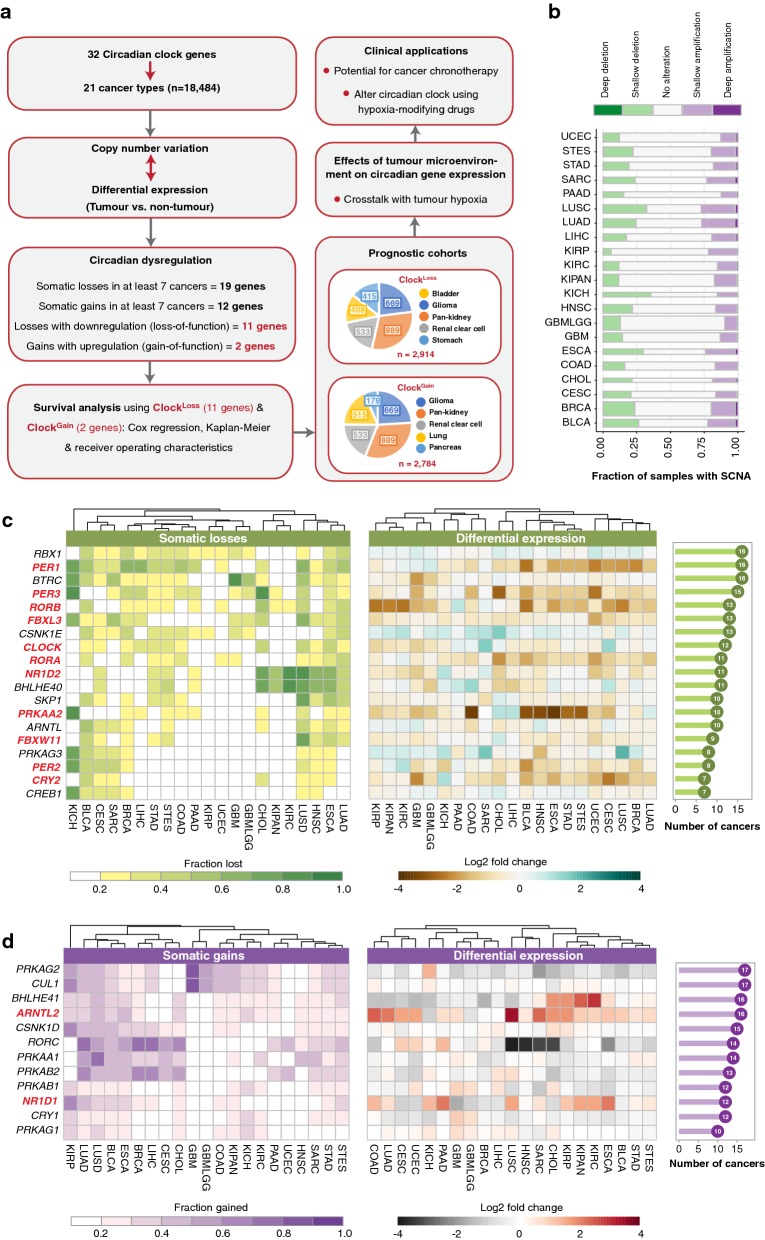
Methods: In an integrated approach uniting genomic, transcriptomic and clinical data of 21 cancer types (n = 18,484), we interrogated copy number and transcript profiles of 32 circadian clock genes to identify putative loss-of-function (ClockLoss) and gain-of-function (ClockGain) players. Kaplan-Meier, Cox regression and receiver operating characteristic analyses were employed to evaluate the prognostic significance of both gene sets.
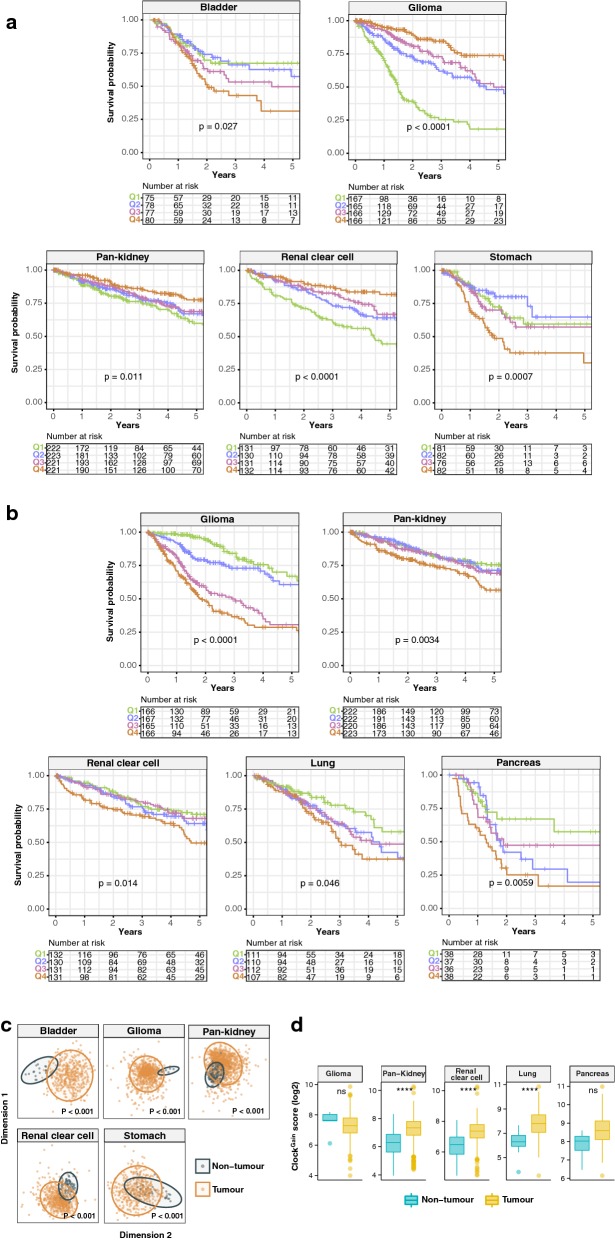
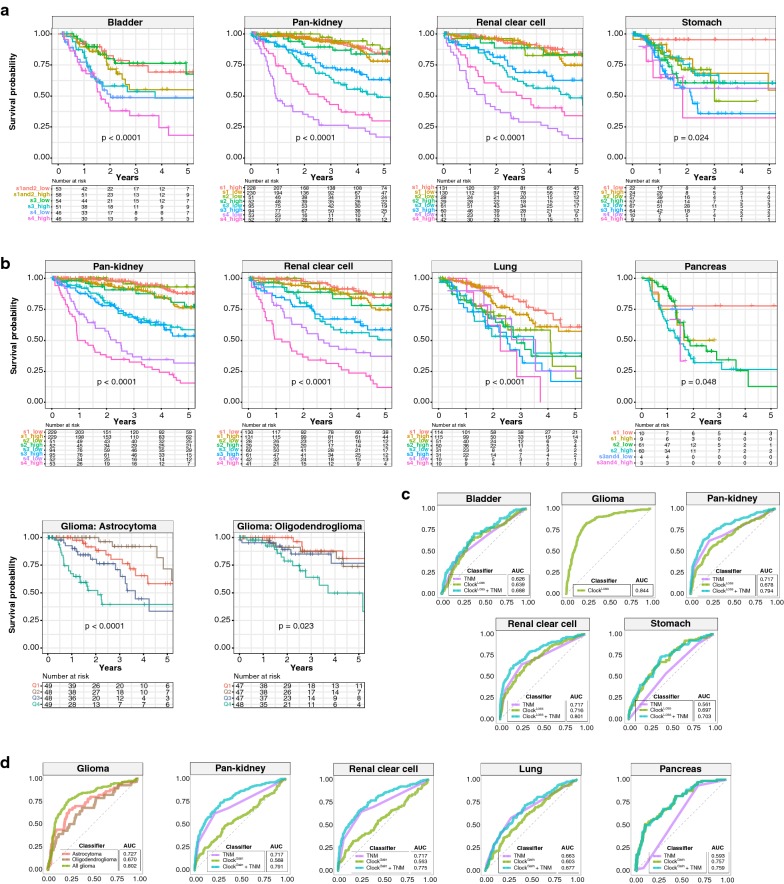
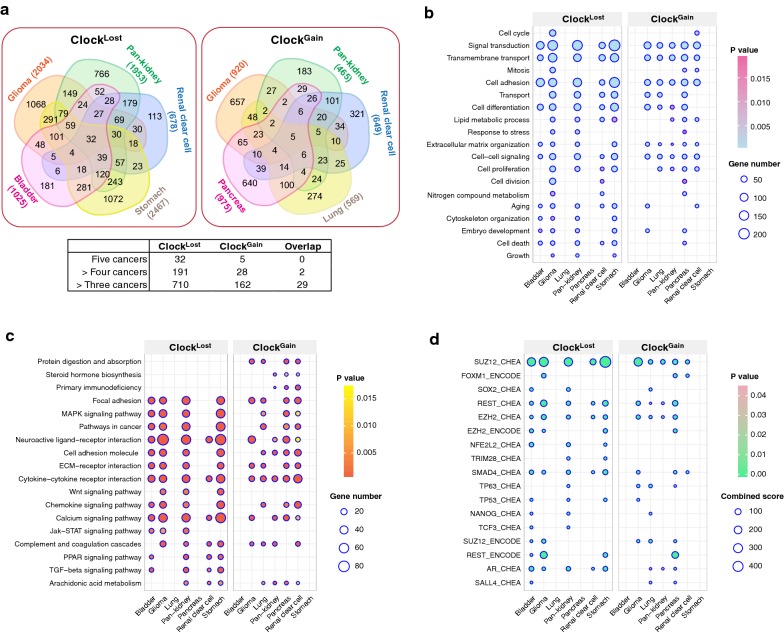
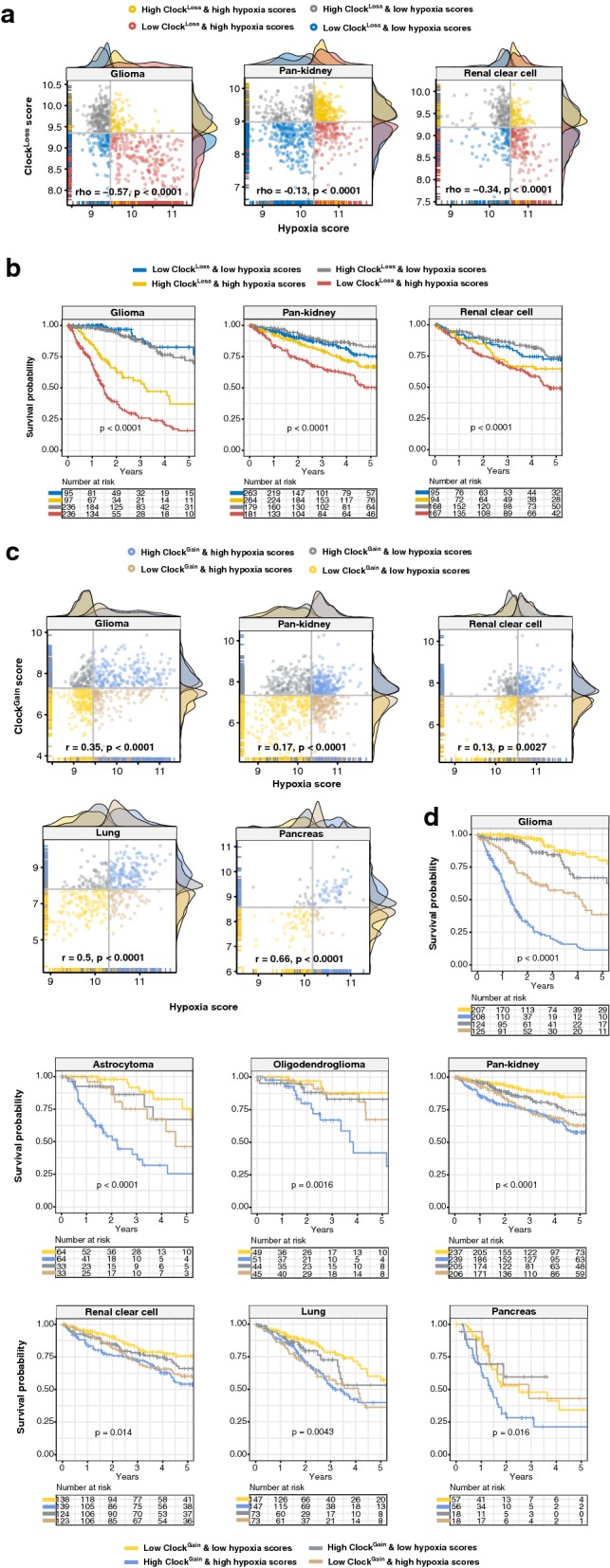
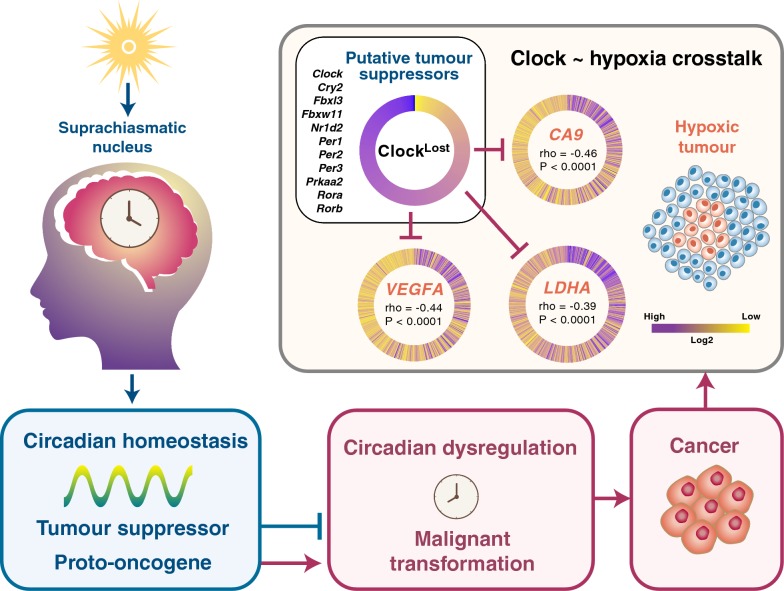
Results: ClockLoss and ClockGain were associated with tumour-suppressing and tumour-promoting roles respectively. Downregulation of ClockLoss genes resulted in significantly higher mortality rates in five cancer cohorts (n = 2914): bladder (P = 0.027), glioma (P < 0.0001), pan-kidney (P = 0.011), clear cell renal cell (P < 0.0001) and stomach (P = 0.0007). In contrast, patients with high expression of oncogenic ClockGain genes had poorer survival outcomes (n = 2784): glioma (P < 0.0001), pan-kidney (P = 0.0034), clear cell renal cell (P = 0.014), lung (P = 0.046) and pancreas (P = 0.0059). Both gene sets were independent of other clinicopathological features to permit further delineation of tumours within the same stage. Circadian reprogramming of tumour genomes resulted in activation of numerous oncogenic pathways including those associated with cancer stem cells, suggesting that the circadian clock may influence self-renewal mechanisms. Within the hypoxic tumour microenvironment, circadian dysregulation is exacerbated by tumour hypoxia in glioma, renal, lung and pancreatic cancers, resulting in additional death risks. Tumour suppressive ClockLoss genes were negatively correlated with hypoxia inducible factor-1A targets in glioma patients, providing a novel framework for investigating the hypoxia-clock signalling axis.
Conclusions: Loss of timekeeping fidelity promotes tumour progression and influences clinical outcomes. ClockLoss and ClockGain may offer novel druggable targets for improving patient prognosis. Both gene sets can be used for patient stratification in adjuvant chronotherapy treatment. Emerging interactions between the circadian clock and hypoxia may be harnessed to achieve therapeutic advantage using hypoxia-modifying compounds in combination with first-line treatments.
Keywords: Circadian clock; Gain-of-function; Glioma; Hypoxia; Loss-of-function; Oncogene; Pan-cancer; Renal cancer; Tumour suppressor.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
An immunoevasive strategy through clinically-relevant pan-cancer genomic and transcriptomic alterations of JAK-STAT signaling components.Mol Med. 2019 Nov 4;25(1):46. doi: 10.1186/s10020-019-0114-1. Mol Med. 2019. PMID: 31684858 Free PMC article.
-
Aberrations in Notch-Hedgehog signalling reveal cancer stem cells harbouring conserved oncogenic properties associated with hypoxia and immunoevasion.Br J Cancer. 2019 Oct;121(8):666-678. doi: 10.1038/s41416-019-0572-9. Epub 2019 Sep 16. Br J Cancer. 2019. PMID: 31523055 Free PMC article.
-
Transcriptional landscape of DNA repair genes underpins a pan-cancer prognostic signature associated with cell cycle dysregulation and tumor hypoxia.DNA Repair (Amst). 2019 Jun;78:142-153. doi: 10.1016/j.dnarep.2019.04.008. Epub 2019 Apr 24. DNA Repair (Amst). 2019. PMID: 31054516
-
The circadian clock and the hypoxic response pathway in kidney cancer.Tumour Biol. 2014 Jan;35(1):1-7. doi: 10.1007/s13277-013-1076-5. Epub 2013 Aug 14. Tumour Biol. 2014. PMID: 23943393 Review.
-
Circadian Clocks and Cancer: Timekeeping Governs Cellular Metabolism.Trends Endocrinol Metab. 2019 Jul;30(7):445-458. doi: 10.1016/j.tem.2019.05.001. Epub 2019 May 30. Trends Endocrinol Metab. 2019. PMID: 31155396 Free PMC article. Review.
Cited by
-
An integrative pan-cancer investigation reveals common genetic and transcriptional alterations of AMPK pathway genes as important predictors of clinical outcomes across major cancer types.BMC Cancer. 2020 Aug 17;20(1):773. doi: 10.1186/s12885-020-07286-2. BMC Cancer. 2020. PMID: 32807122 Free PMC article.
-
An immunoevasive strategy through clinically-relevant pan-cancer genomic and transcriptomic alterations of JAK-STAT signaling components.Mol Med. 2019 Nov 4;25(1):46. doi: 10.1186/s10020-019-0114-1. Mol Med. 2019. PMID: 31684858 Free PMC article.
-
Insights About Circadian Clock and Molecular Pathogenesis in Gliomas.Front Oncol. 2020 Feb 28;10:199. doi: 10.3389/fonc.2020.00199. eCollection 2020. Front Oncol. 2020. PMID: 32195174 Free PMC article. Review.
-
Application of ensemble clustering and survival tree analysis for identifying prognostic clinicogenomic features in patients with colorectal cancer from the 100,000 Genomes Project.BMC Res Notes. 2021 Oct 2;14(1):385. doi: 10.1186/s13104-021-05789-0. BMC Res Notes. 2021. PMID: 34600575 Free PMC article.
-
Unravelling the link between circadian clock genes and brain tumors: From pathological disruptions to potential therapeutic interventions.Front Pharmacol. 2025 May 27;16:1617713. doi: 10.3389/fphar.2025.1617713. eCollection 2025. Front Pharmacol. 2025. PMID: 40495887 Free PMC article. Review.
References
MeSH terms
LinkOut - more resources
Full Text Sources
