

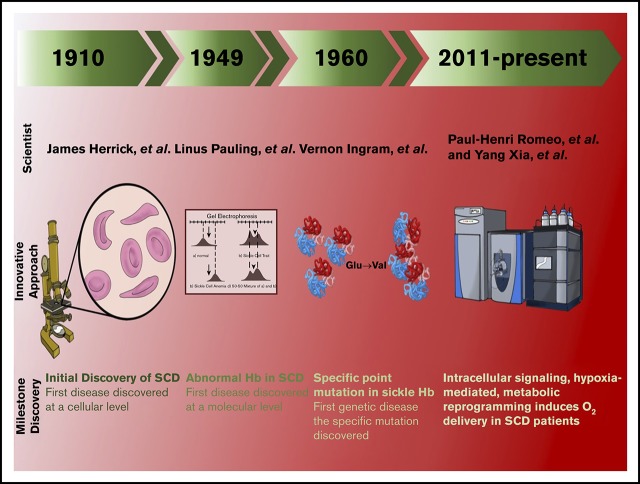
Metabolomic and molecular insights into sickle cell disease and innovative therapies
- PMID: 31015210
- PMCID: PMC6482357
- DOI: 10.1182/bloodadvances.2018030619
Metabolomic and molecular insights into sickle cell disease and innovative therapies
Abstract
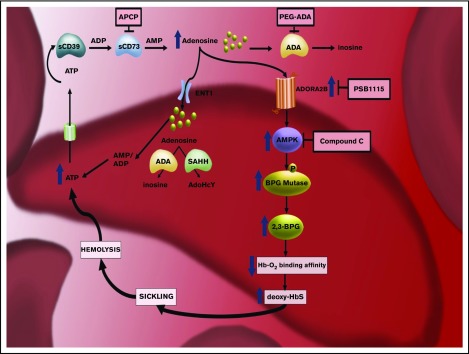
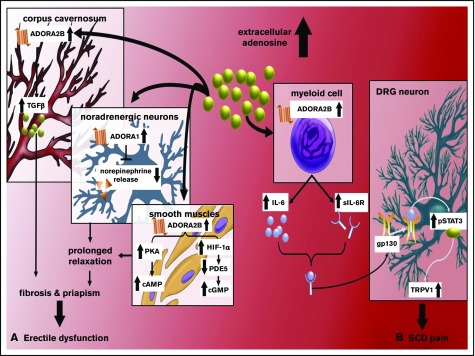
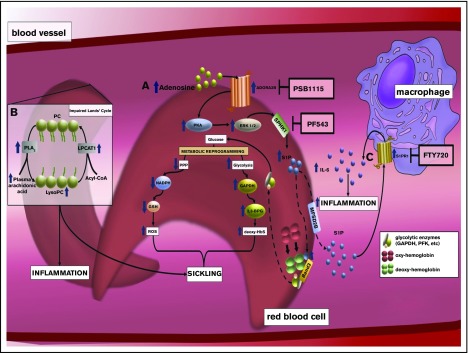
Sickle cell disease (SCD) is an autosomal-recessive hemolytic disorder with high morbidity and mortality. The pathophysiology of SCD is characterized by the polymerization of deoxygenated intracellular sickle hemoglobin, which causes the sickling of erythrocytes. The recent development of metabolomics, the newest member of the "omics" family, has provided a powerful new research strategy to accurately measure functional phenotypes that are the net result of genomic, transcriptomic, and proteomic changes. Metabolomics changes respond faster to external stimuli than any other "ome" and are especially appropriate for surveilling the metabolic profile of erythrocytes. In this review, we summarize recent pioneering research that exploited cutting-edge metabolomics and state-of-the-art isotopically labeled nutrient flux analysis to monitor and trace intracellular metabolism in SCD mice and humans. Genetic, structural, biochemical, and molecular studies in mice and humans demonstrate unrecognized intracellular signaling pathways, including purinergic and sphingolipid signaling networks that promote hypoxic metabolic reprogramming by channeling glucose metabolism to glycolysis via the pentose phosphate pathway. In turn, this hypoxic metabolic reprogramming induces 2,3-bisphosphoglycerate production, deoxygenation of sickle hemoglobin, polymerization, and sickling. Additionally, we review the detrimental role of an impaired Lands' cycle, which contributes to sickling, inflammation, and disease progression. Thus, metabolomic profiling allows us to identify the pathological role of adenosine signaling and S1P-mediated erythrocyte hypoxic metabolic reprogramming and hypoxia-induced impaired Lands' cycle in SCD. These findings further reveal that the inhibition of adenosine and S1P signaling cascade and the restoration of an imbalanced Lands' cycle have potent preclinical efficacy in counteracting sickling, inflammation, and disease progression.
© 2019 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- Pauling L, Itano HA, Singer SJ, Wells IC. Sickle cell anemia a molecular disease. Science. 1949;110(2865):543-548. - PubMed
-
- Wells IC, Itano HA. Ratio of sickle-cell anemia hemoglobin to normal hemoglobin in sicklemics. J Biol Chem. 1951;188(1):65-74. - PubMed
-
- Eaton WA. Linus Pauling and sickle cell disease. Biophys Chem. 2003;100(1-3):109-116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
