

A mutation in CaV2.1 linked to a severe neurodevelopmental disorder impairs channel gating
- PMID: 31015257
- PMCID: PMC6571999
- DOI: 10.1085/jgp.201812237
A mutation in CaV2.1 linked to a severe neurodevelopmental disorder impairs channel gating
Abstract
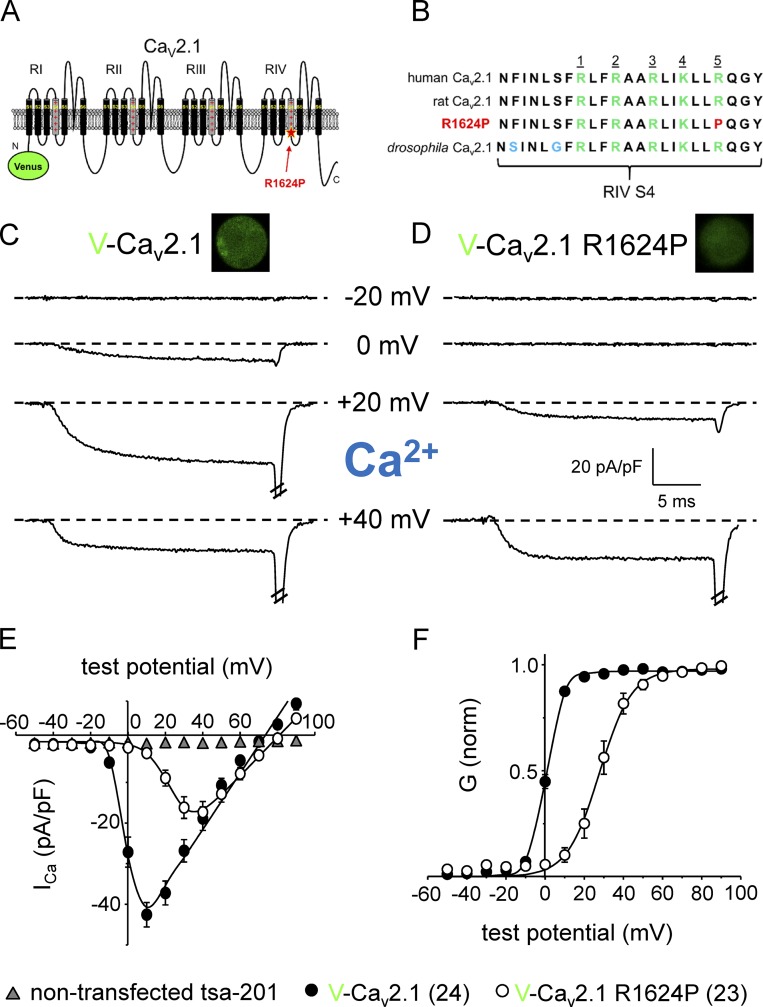
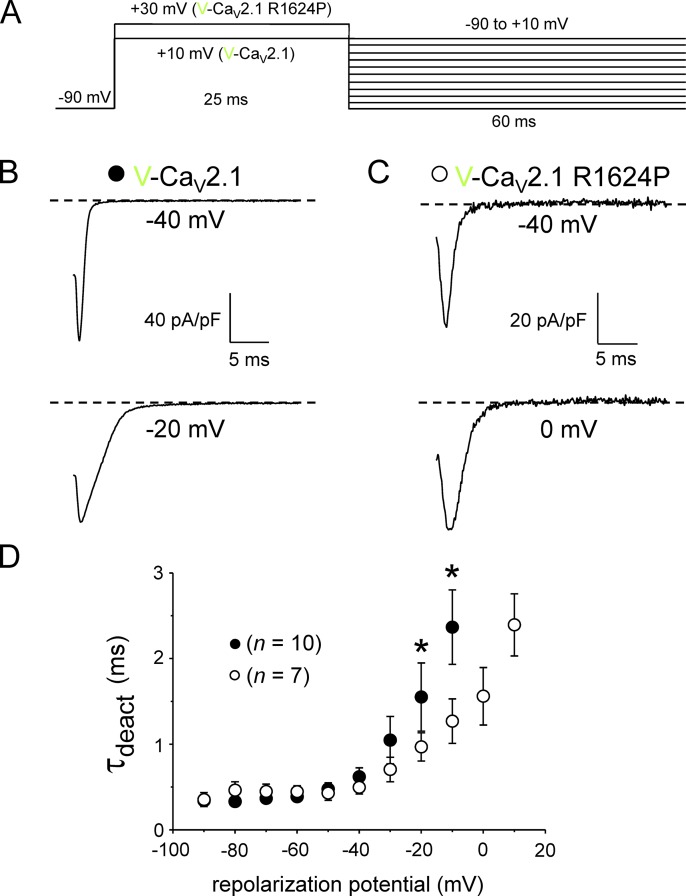
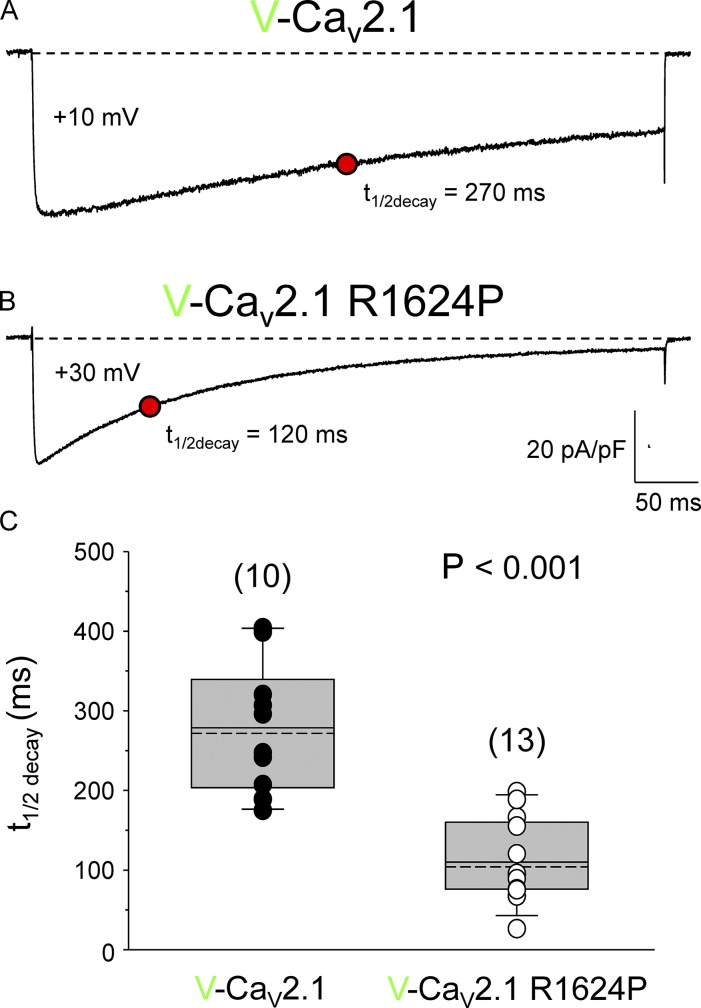
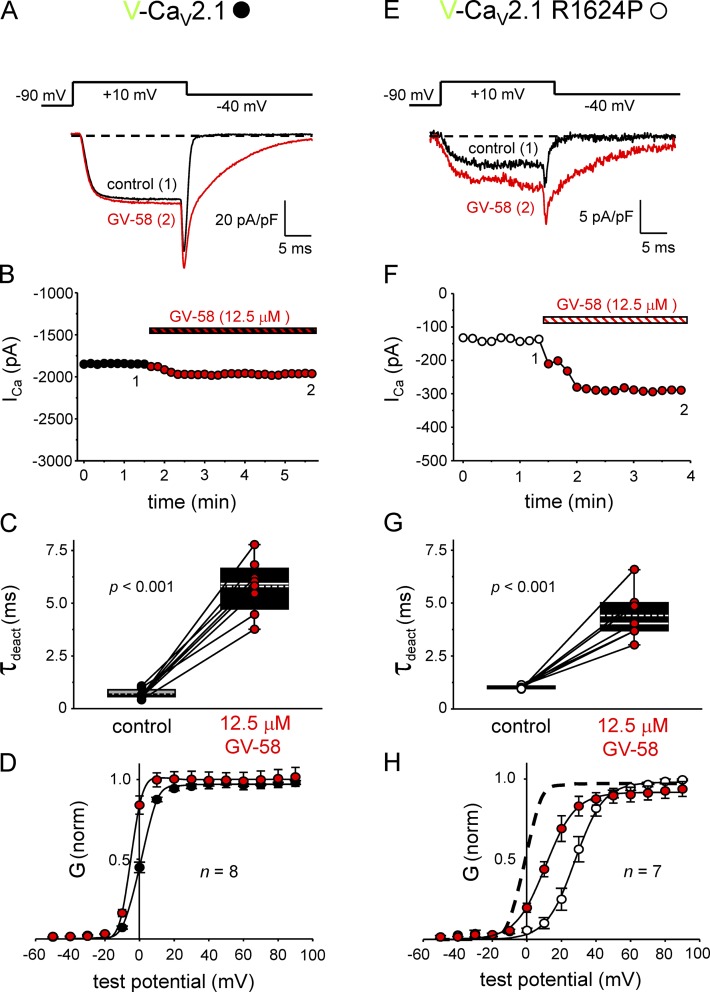
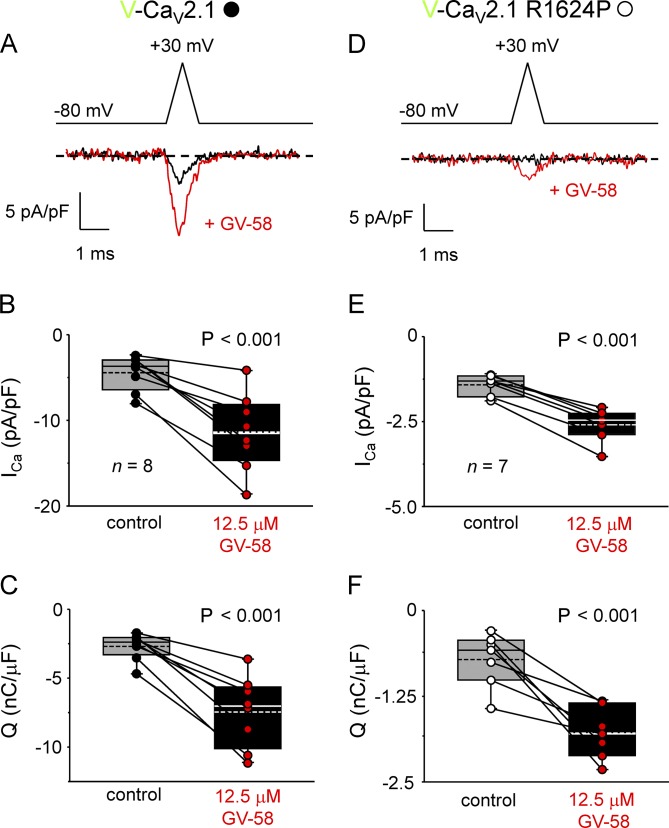
Ca2+ flux into axon terminals via P-/Q-type CaV2.1 channels is the trigger for neurotransmitter vesicle release at neuromuscular junctions (NMJs) and many central synapses. Recently, an arginine to proline substitution (R1673P) in the S4 voltage-sensing helix of the fourth membrane-bound repeat of CaV2.1 was linked to a severe neurological disorder characterized by generalized hypotonia, ataxia, cerebellar atrophy, and global developmental delay. The R1673P mutation was proposed to cause a gain of function in CaV2.1 leading to neuronal Ca2+ toxicity based on the ability of the mutant channel to rescue the photoreceptor response in CaV2.1-deficient Drosophila cacophony larvae. Here, we show that the corresponding mutation in rat CaV2.1 (R1624P) causes a profound loss of channel function; voltage-clamp analysis of tsA-201 cells expressing this mutant channel revealed an ∼25-mV depolarizing shift in the voltage dependence of activation. This alteration in activation implies that a significant fraction of CaV2.1 channels resident in presynaptic terminals are unlikely to open in response to an action potential, thereby increasing the probability of synaptic failure at both NMJs and central synapses. Indeed, the mutant channel supported only minimal Ca2+ flux in response to an action potential-like waveform. Application of GV-58, a compound previously shown to stabilize the open state of wild-type CaV2.1 channels, partially restored Ca2+ current by shifting mutant activation to more hyperpolarizing potentials and slowing deactivation. Consequently, GV-58 also rescued a portion of Ca2+ flux during action potential-like stimuli. Thus, our data raise the possibility that therapeutic agents that increase channel open probability or prolong action potential duration may be effective in combatting this and other severe neurodevelopmental disorders caused by loss-of-function mutations in CaV2.1.
© 2019 Tyagi et al.
Figures

References
-
- Bahamonde M.I., Serra S.A., Drechsel O., Rahman R., Marcé-Grau A., Prieto M., Ossowski S., Macaya A., and Fernández-Fernández J.M.. 2015. A single amino acid deletion (ΔF1502) in the S6 segment of CaV2.1 domain III associated with congenital ataxia increases channel activity and promotes Ca2+ influx. PLoS One. 10:e0146035 10.1371/journal.pone.0146035 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
