

Tracing the origin and evolution of pseudokinases across the tree of life
- PMID: 31015289
- PMCID: PMC6997932
- DOI: 10.1126/scisignal.aav3810
Tracing the origin and evolution of pseudokinases across the tree of life
Abstract
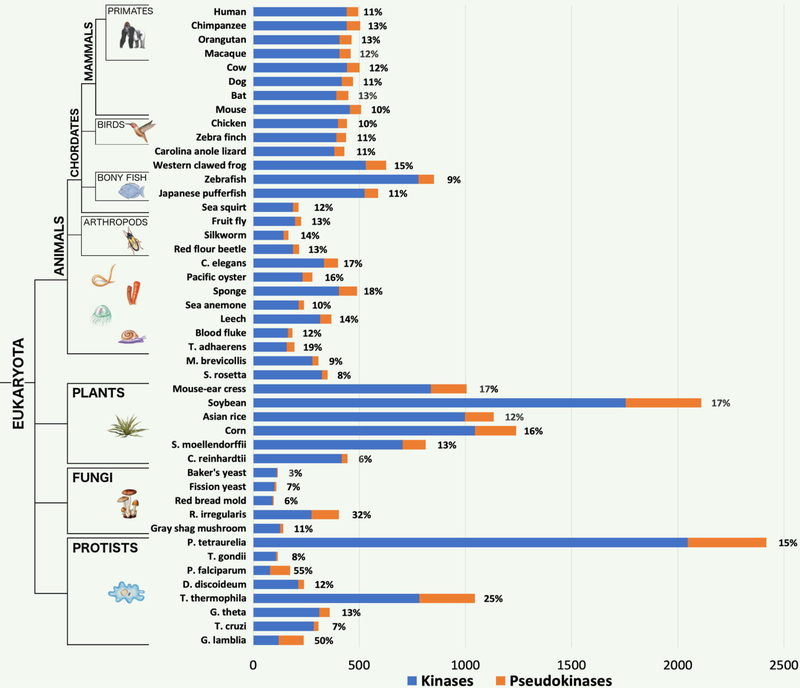
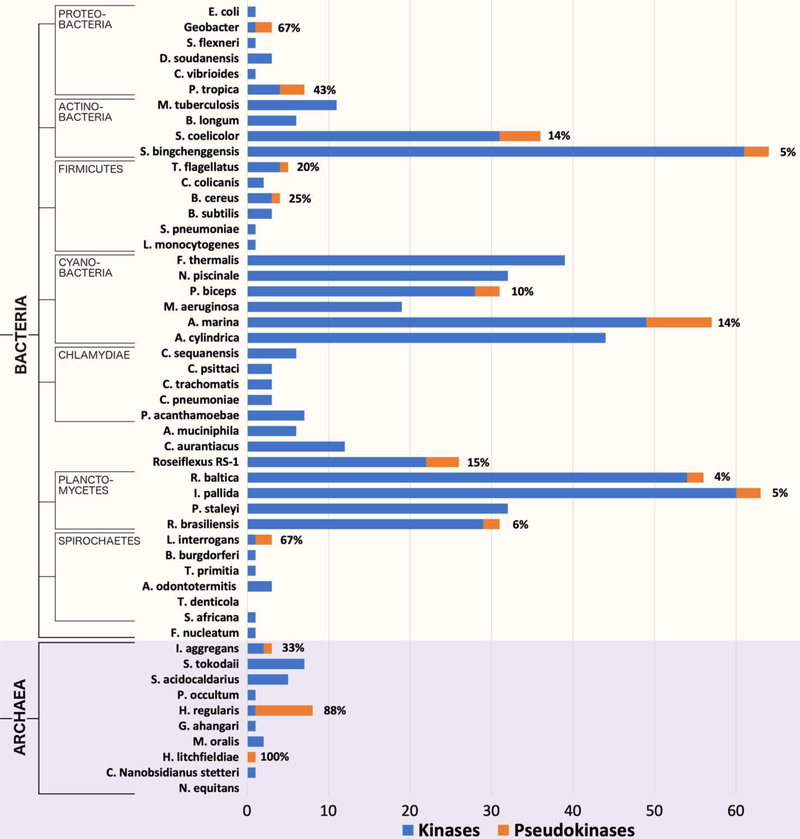
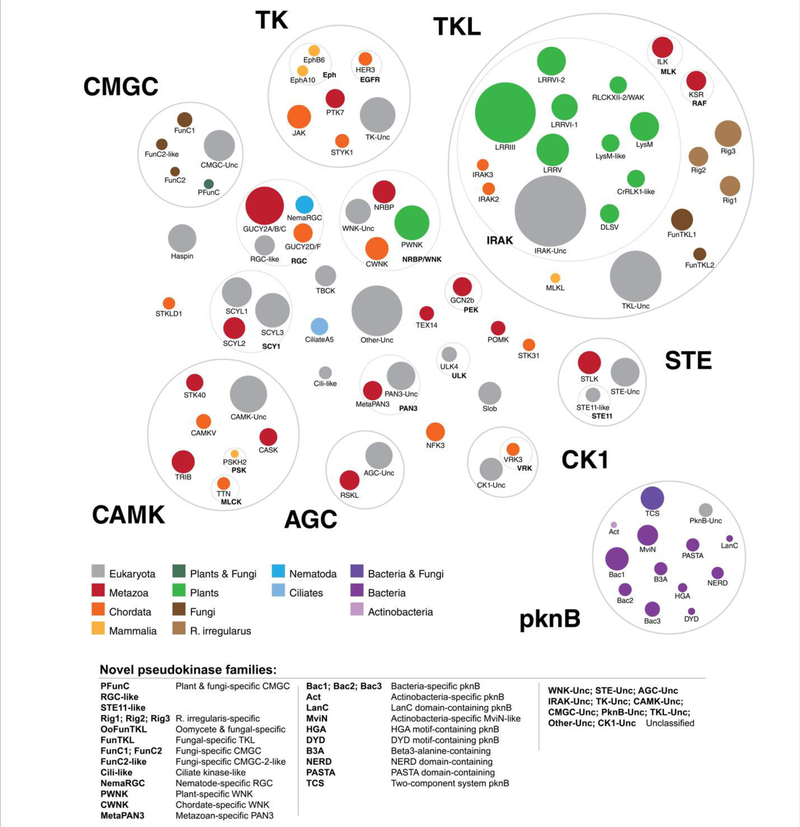
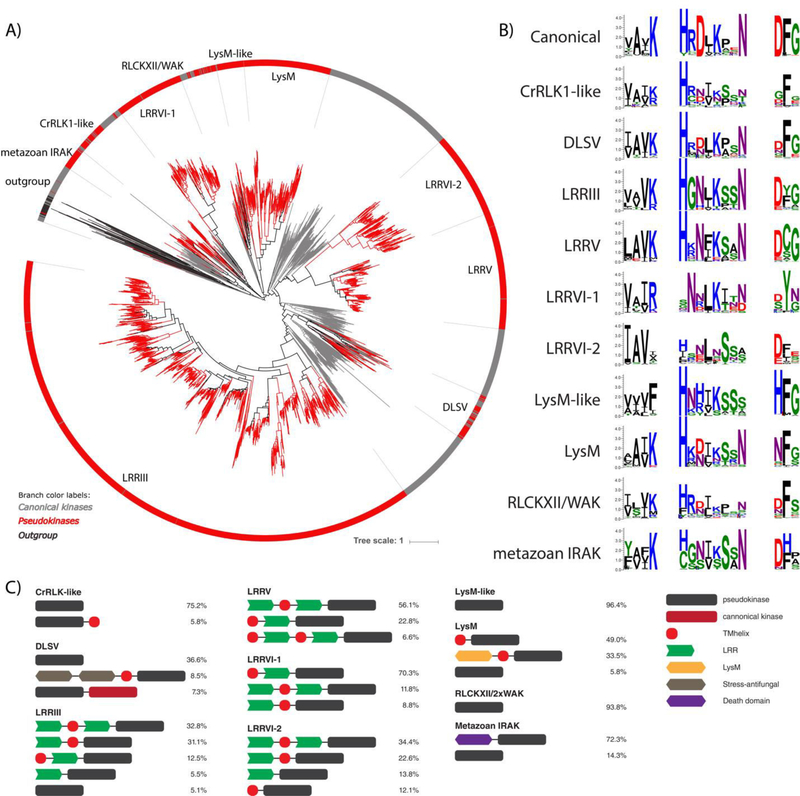
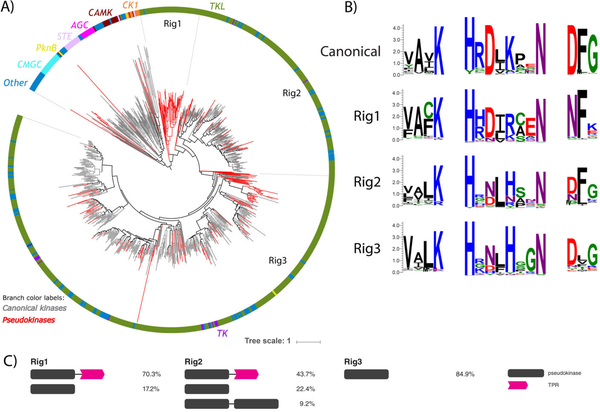
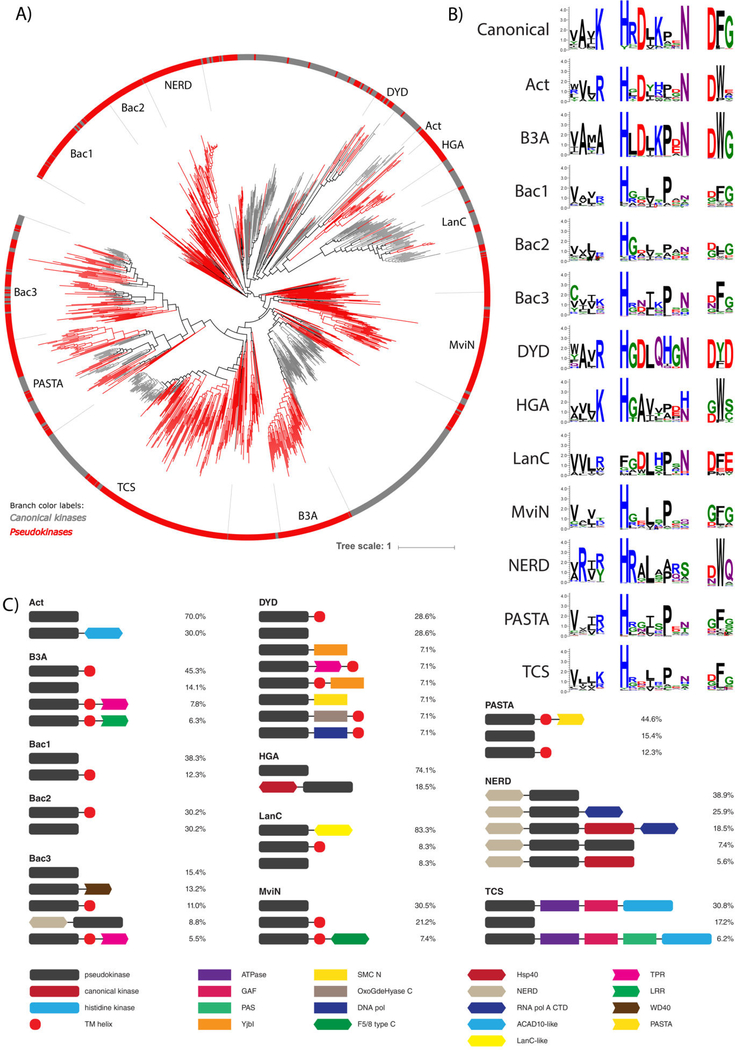
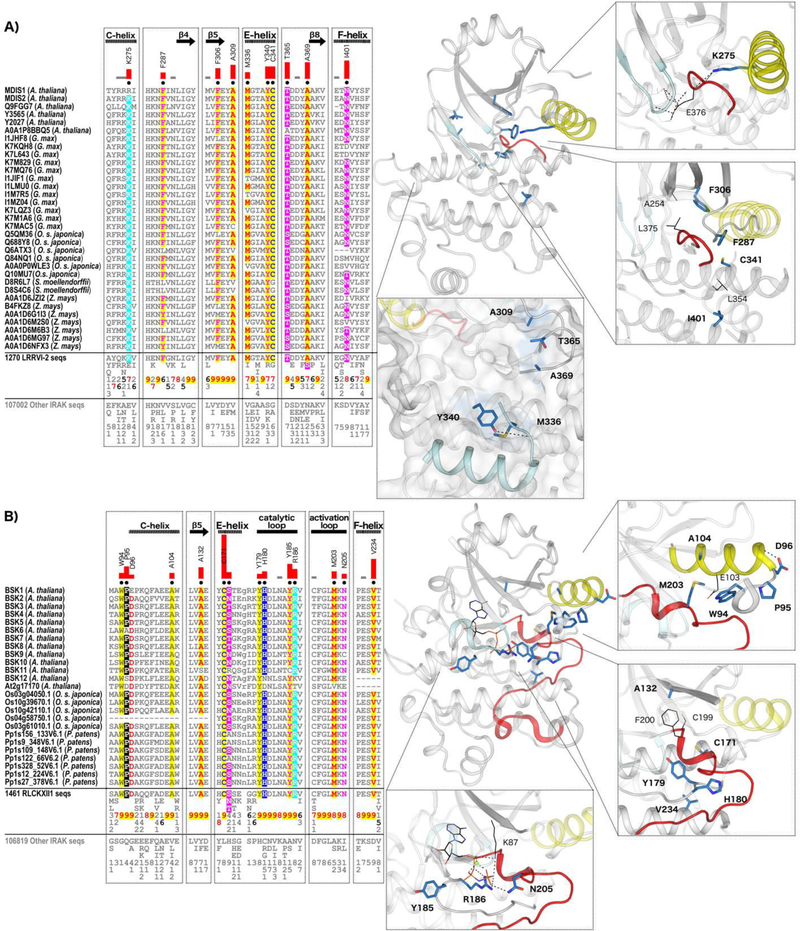
Protein phosphorylation by eukaryotic protein kinases (ePKs) is a fundamental mechanism of cell signaling in all organisms. In model vertebrates, ~10% of ePKs are classified as pseudokinases, which have amino acid changes within the catalytic machinery of the kinase domain that distinguish them from their canonical kinase counterparts. However, pseudokinases still regulate various signaling pathways, usually doing so in the absence of their own catalytic output. To investigate the prevalence, evolutionary relationships, and biological diversity of these pseudoenzymes, we performed a comprehensive analysis of putative pseudokinase sequences in available eukaryotic, bacterial, and archaeal proteomes. We found that pseudokinases are present across all domains of life, and we classified nearly 30,000 eukaryotic, 1500 bacterial, and 20 archaeal pseudokinase sequences into 86 pseudokinase families, including ~30 families that were previously unknown. We uncovered a rich variety of pseudokinases with notable expansions not only in animals but also in plants, fungi, and bacteria, where pseudokinases have previously received cursory attention. These expansions are accompanied by domain shuffling, which suggests roles for pseudokinases in plant innate immunity, plant-fungal interactions, and bacterial signaling. Mechanistically, the ancestral kinase fold has diverged in many distinct ways through the enrichment of unique sequence motifs to generate new families of pseudokinases in which the kinase domain is repurposed for noncanonical nucleotide binding or to stabilize unique, inactive kinase conformations. We further provide a collection of annotated pseudokinase sequences in the Protein Kinase Ontology (ProKinO) as a new mineable resource for the signaling community.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Cohen P, The origins of protein phosphorylation. Nat Cell Biol 4, E127–130 (2002). - PubMed
-
- Knighton DR et al., Crystal structure of the catalytic subunit of cyclic adenosine monophosphate-dependent protein kinase. Science 253, 407–414 (1991). - PubMed
-
- Knighton DR et al., Structure of a peptide inhibitor bound to the catalytic subunit of cyclic adenosine monophosphate-dependent protein kinase. Science 253, 414–420 (1991). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
