

Quantitative Analyses Reveal Novel Roles for N- Glycosylation in a Major Enteric Bacterial Pathogen
- PMID: 31015322
- PMCID: PMC6478998
- DOI: 10.1128/mBio.00297-19
Quantitative Analyses Reveal Novel Roles for N- Glycosylation in a Major Enteric Bacterial Pathogen
Abstract
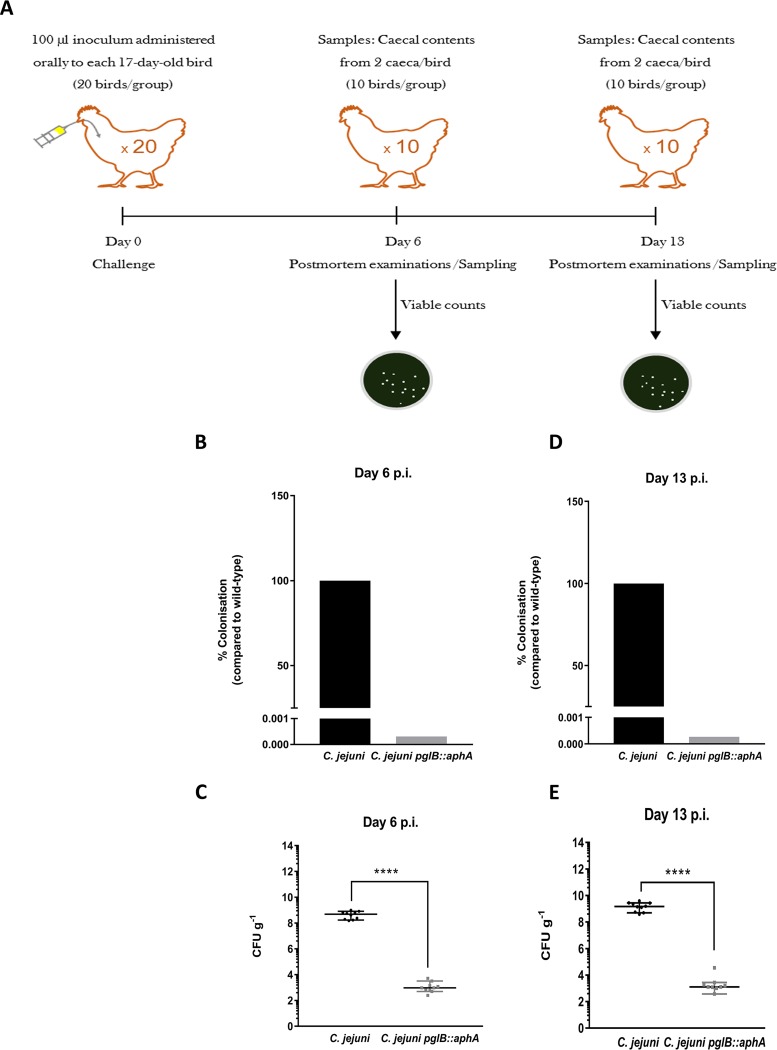
In eukaryotes, glycosylation plays a role in proteome stability, protein quality control, and modulating protein function; however, similar studies in bacteria are lacking. Here, we investigate the roles of general protein glycosylation systems in bacteria using the enteropathogen Campylobacter jejuni as a well-defined example. By using a quantitative proteomic strategy, we were able to monitor changes in the C. jejuni proteome when glycosylation is disrupted. We demonstrate that in C. jejuni, N-glycosylation is essential to maintain proteome stability and protein quality control. These findings guided us to investigate the role of N-glycosylation in modulating bacterial cellular activities. In glycosylation-deficient C. jejuni, the multidrug efflux pump and electron transport pathways were significantly impaired. We demonstrate that in vivo, fully glycosylation-deficient C. jejuni bacteria were unable to colonize its natural avian host. These results provide the first evidence of a link between proteome stability and complex functions via a bacterial general glycosylation system.IMPORTANCE Advances in genomics and mass spectrometry have revealed several types of glycosylation systems in bacteria. However, why bacterial proteins are modified remains poorly defined. Here, we investigated the role of general N-linked glycosylation in a major food poisoning bacterium, Campylobacter jejuni The aim of this study is to delineate the direct and indirect effects caused by disrupting this posttranslational modification. To achieve this, we employed a quantitative proteomic strategy to monitor alterations in the C. jejuni proteome. Our quantitative proteomic results linked general protein N-glycosylation to maintaining proteome stability. Functional analyses revealed novel roles for bacterial N-glycosylation in modulating multidrug efflux pump, enhancing nitrate reduction activity, and promoting host-microbe interaction. This work provides insights on the importance of general glycosylation in proteins in maintaining bacterial physiology, thus expanding our knowledge of the emergence of posttranslational modification in bacteria.
Keywords: glycosylation; host-microbe interaction; microbial physiology; pathogenesis; proteomics.
Copyright © 2019 Abouelhadid et al.
Figures
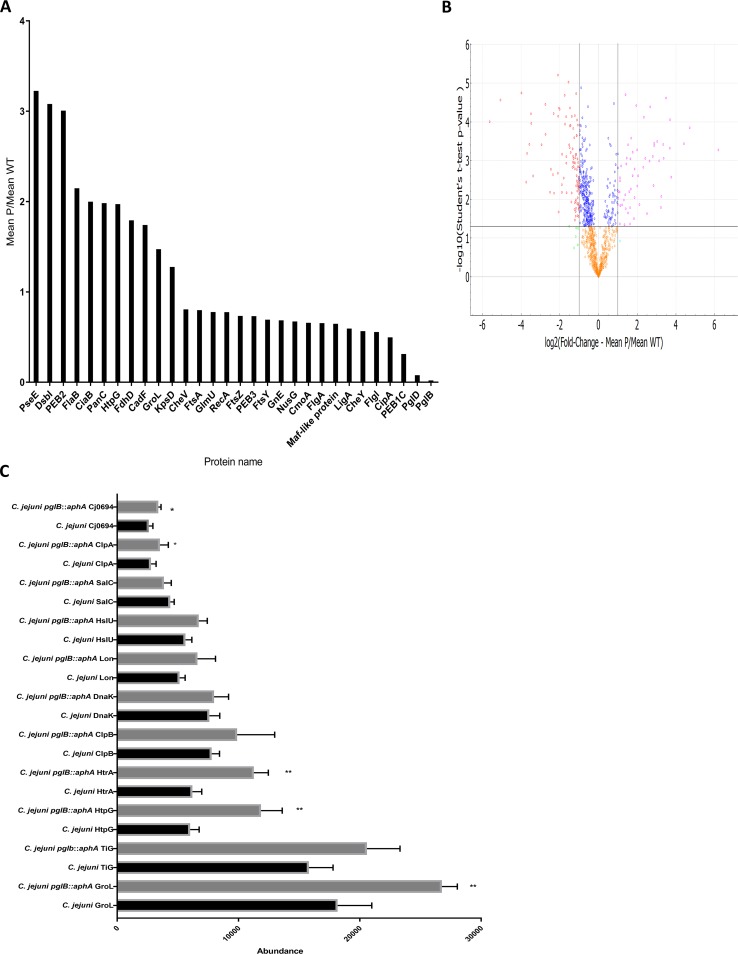
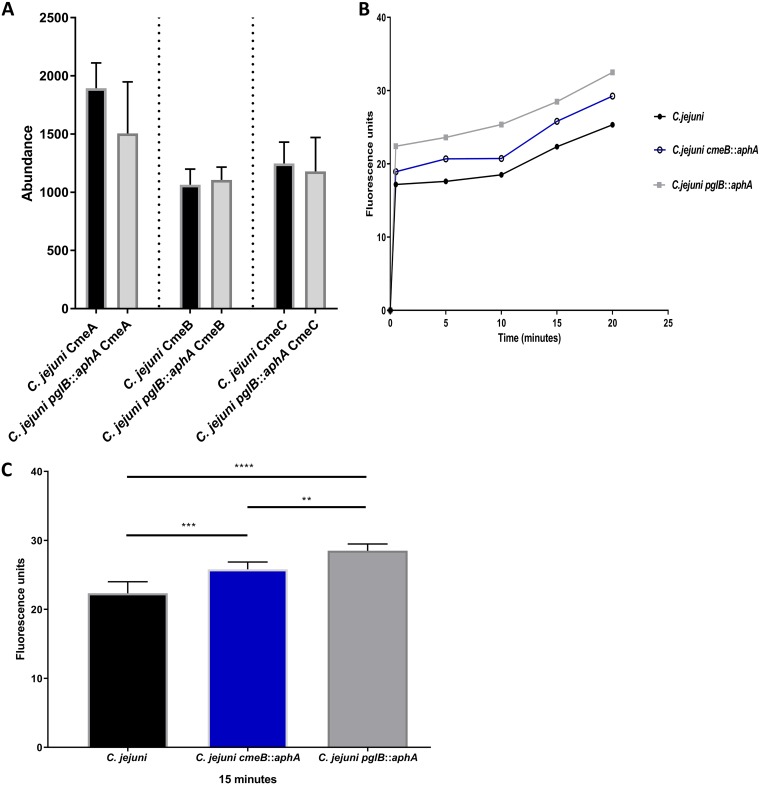
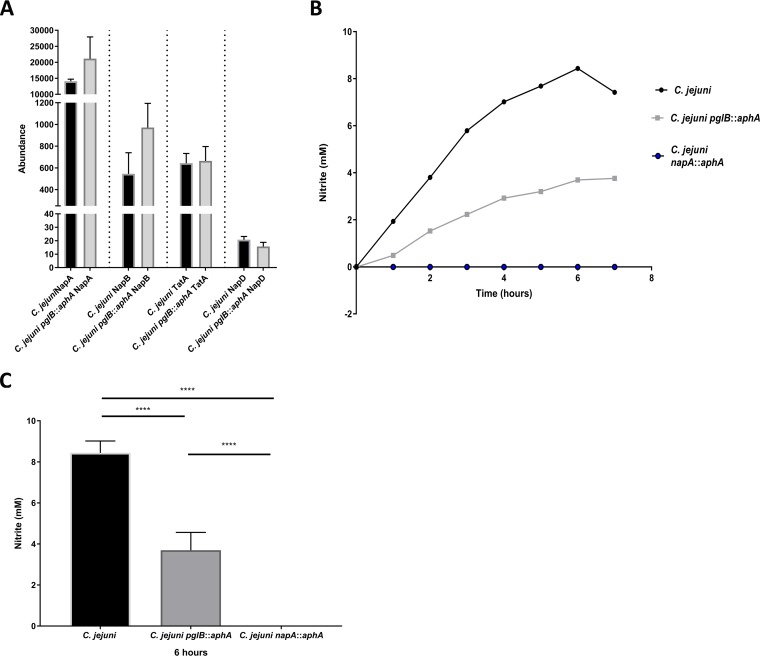
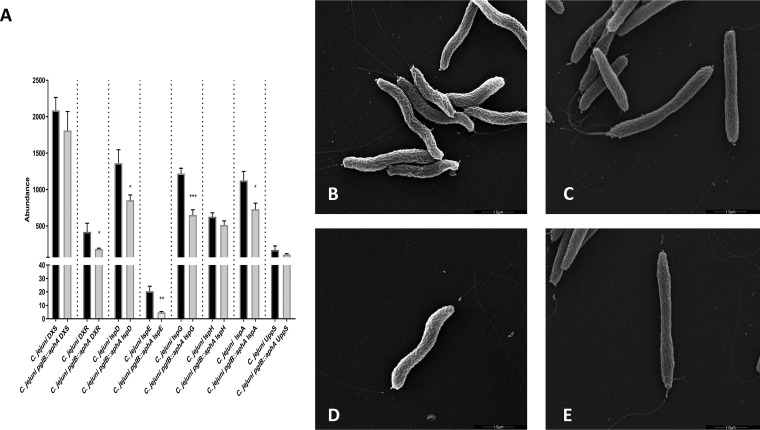
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
