

Stable and Highly Immunogenic MicroRNA-Targeted Single-Dose Live Attenuated Vaccine Candidate against Tick-Borne Encephalitis Constructed Using Genetic Backbone of Langat Virus
- PMID: 31015334
- PMCID: PMC6479010
- DOI: 10.1128/mBio.02904-18
Stable and Highly Immunogenic MicroRNA-Targeted Single-Dose Live Attenuated Vaccine Candidate against Tick-Borne Encephalitis Constructed Using Genetic Backbone of Langat Virus
Abstract
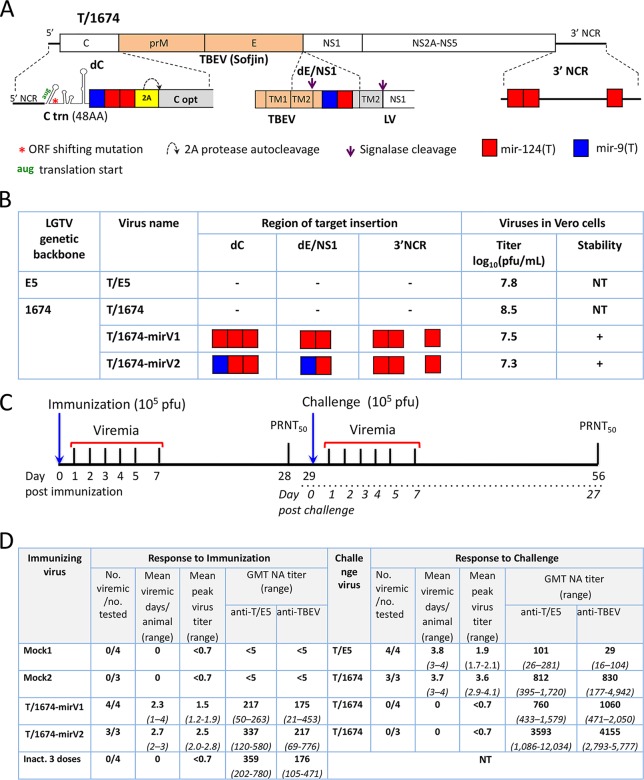
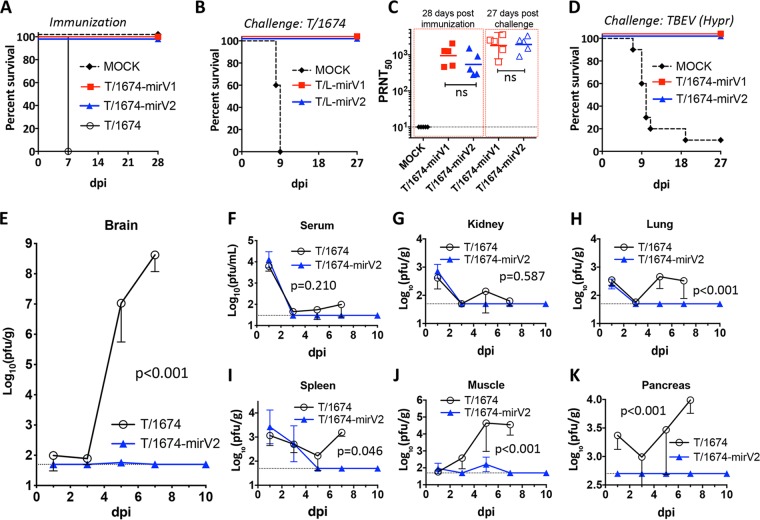
Tick-borne encephalitis virus (TBEV), a member of the genus Flavivirus, is one of the most medically important tick-borne pathogens of the Old World. Despite decades of active research, attempts to develop of a live attenuated virus (LAV) vaccine against TBEV with acceptable safety and immunogenicity characteristics have not been successful. To overcome this impasse, we generated a chimeric TBEV that was highly immunogenic in nonhuman primates (NHPs). The chimeric virus contains the prM/E genes of TBEV, which are expressed in the genetic background of an antigenically closely related, but less pathogenic member of the TBEV complex-Langat virus (LGTV), strain T-1674. The neurovirulence of this chimeric virus was subsequently controlled by robust targeting of the viral genome with multiple copies of central nervous system-enriched microRNAs (miRNAs). This miRNA-targeted T/1674-mirV2 virus was highly stable in Vero cells and was not pathogenic in various mouse models of infection or in NHPs. Importantly, in NHPs, a single dose of the T/1674-mirV2 virus induced TBEV-specific neutralizing antibody (NA) levels comparable to those seen with a three-dose regimen of an inactivated TBEV vaccine, currently available in Europe. Moreover, our vaccine candidate provided complete protection against a stringent wild-type TBEV challenge in mice and against challenge with a parental (not miRNA-targeted) chimeric TBEV/LGTV in NHPs. Thus, this highly attenuated and immunogenic T/1674-mirV2 virus is a promising LAV vaccine candidate against TBEV and warrants further preclinical evaluation of its neurovirulence in NHPs prior to entering clinical trials in humans.IMPORTANCE Tick-borne encephalitis virus (TBEV) is one of the most medically important tick-borne pathogens of the Old World. Despite decades of active research, efforts to develop of TBEV live attenuated virus (LAV) vaccines with acceptable safety and immunogenicity characteristics have not been successful. Here we report the development and evaluation of a highly attenuated and immunogenic microRNA-targeted TBEV LAV.
Keywords: Langat virus; chimeric virus; live attenuated virus vaccine; microRNA; tick-borne encephalitis virus.
Figures
Similar articles
-
Tick-borne Langat/mosquito-borne dengue flavivirus chimera, a candidate live attenuated vaccine for protection against disease caused by members of the tick-borne encephalitis virus complex: evaluation in rhesus monkeys and in mosquitoes.J Virol. 2001 Sep;75(17):8259-67. doi: 10.1128/jvi.75.17.8259-8267.2001. J Virol. 2001. PMID: 11483771 Free PMC article.
-
Evaluation of the Langat/dengue 4 chimeric virus as a live attenuated tick-borne encephalitis vaccine for safety and immunogenicity in healthy adult volunteers.Vaccine. 2008 Feb 13;26(7):882-90. doi: 10.1016/j.vaccine.2007.12.015. Epub 2008 Jan 4. Vaccine. 2008. PMID: 18207289 Free PMC article. Clinical Trial.
-
Recombinant chimeric Japanese encephalitis virus/tick-borne encephalitis virus is attenuated and protective in mice.Vaccine. 2014 Feb 12;32(8):949-56. doi: 10.1016/j.vaccine.2013.12.050. Epub 2014 Jan 4. Vaccine. 2014. PMID: 24394443
-
Immunogenicity against Far Eastern and Siberian subtypes of tick-borne encephalitis (TBE) virus elicited by the currently available vaccines based on the European subtype: systematic review and meta-analysis.Hum Vaccin Immunother. 2014;10(10):2819-33. doi: 10.4161/hv.29984. Hum Vaccin Immunother. 2014. PMID: 25483679 Free PMC article.
-
Tick-borne encephalitis.Antiviral Res. 2003 Jan;57(1-2):129-46. doi: 10.1016/s0166-3542(02)00206-1. Antiviral Res. 2003. PMID: 12615309 Review.
Cited by
-
Novel Approach for Insertion of Heterologous Sequences into Full-Length ZIKV Genome Results in Superior Level of Gene Expression and Insert Stability.Viruses. 2020 Jan 3;12(1):61. doi: 10.3390/v12010061. Viruses. 2020. PMID: 31947825 Free PMC article.
-
Tick-Borne Encephalitis (TBE): From Tick to Pathology.J Clin Med. 2023 Oct 30;12(21):6859. doi: 10.3390/jcm12216859. J Clin Med. 2023. PMID: 37959323 Free PMC article. Review.
-
CpG content in the Zika virus genome affects infection phenotypes in the adult brain and fetal lymph nodes.Front Immunol. 2022 Aug 2;13:943481. doi: 10.3389/fimmu.2022.943481. eCollection 2022. Front Immunol. 2022. PMID: 35983032 Free PMC article.
-
Editorial: Immunological aspects of emerging and re-emerging zoonoses.Front Immunol. 2024 Mar 6;15:1392382. doi: 10.3389/fimmu.2024.1392382. eCollection 2024. Front Immunol. 2024. PMID: 38510255 Free PMC article. No abstract available.
-
Tick-Borne Encephalitis Virus: A Quest for Better Vaccines against a Virus on the Rise.Vaccines (Basel). 2020 Aug 12;8(3):451. doi: 10.3390/vaccines8030451. Vaccines (Basel). 2020. PMID: 32806696 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
