

Ceramides bind VDAC2 to trigger mitochondrial apoptosis
- PMID: 31015432
- PMCID: PMC6478893
- DOI: 10.1038/s41467-019-09654-4
Ceramides bind VDAC2 to trigger mitochondrial apoptosis
Abstract
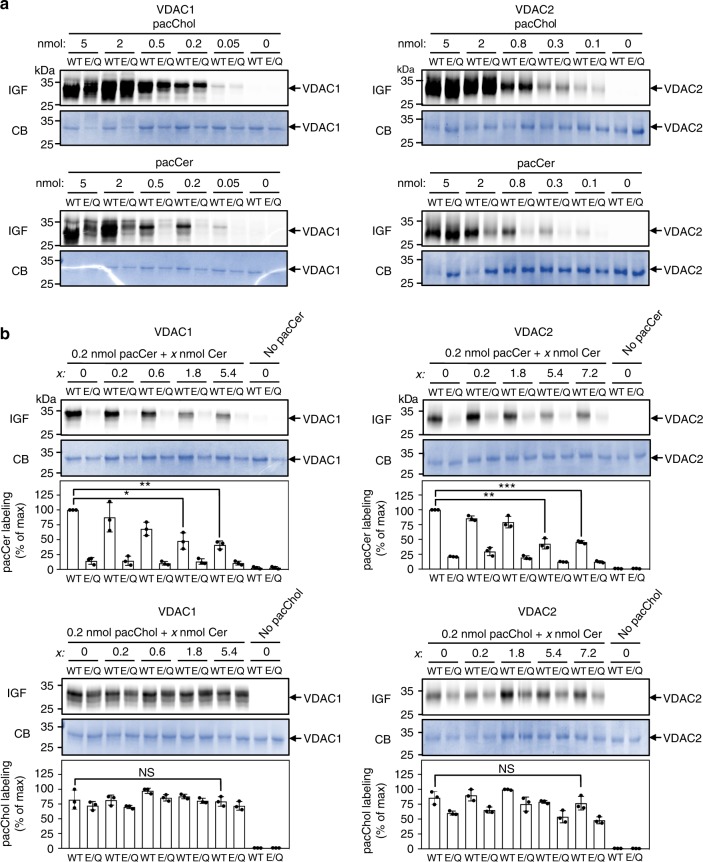
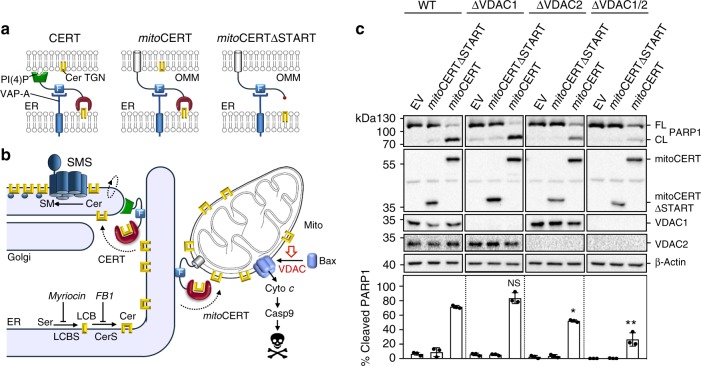
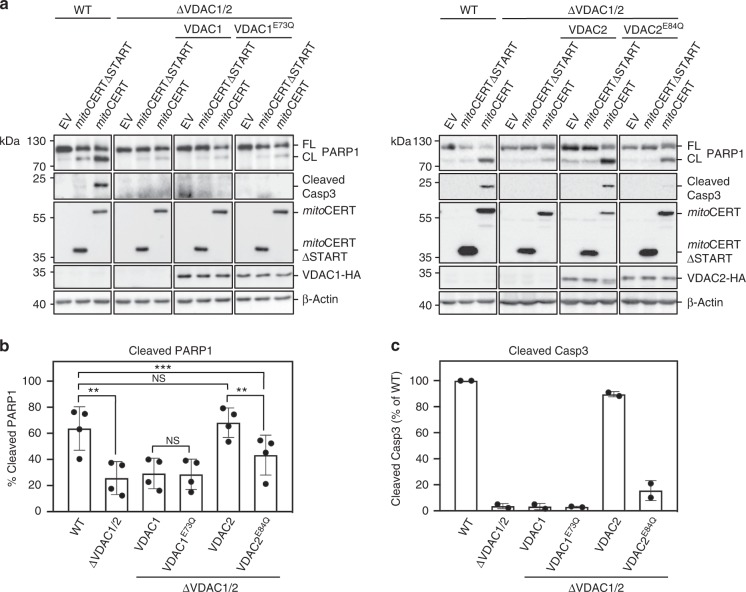
Ceramides draw wide attention as tumor suppressor lipids that act directly on mitochondria to trigger apoptotic cell death. However, molecular details of the underlying mechanism are largely unknown. Using a photoactivatable ceramide probe, we here identify the voltage-dependent anion channels VDAC1 and VDAC2 as mitochondrial ceramide binding proteins. Coarse-grain molecular dynamics simulations reveal that both channels harbor a ceramide binding site on one side of the barrel wall. This site includes a membrane-buried glutamate that mediates direct contact with the ceramide head group. Substitution or chemical modification of this residue abolishes photolabeling of both channels with the ceramide probe. Unlike VDAC1 removal, loss of VDAC2 or replacing its membrane-facing glutamate with glutamine renders human colon cancer cells largely resistant to ceramide-induced apoptosis. Collectively, our data support a role of VDAC2 as direct effector of ceramide-mediated cell death, providing a molecular framework for how ceramides exert their anti-neoplastic activity.
Conflict of interest statement
The authors declare no competing interests.
Figures
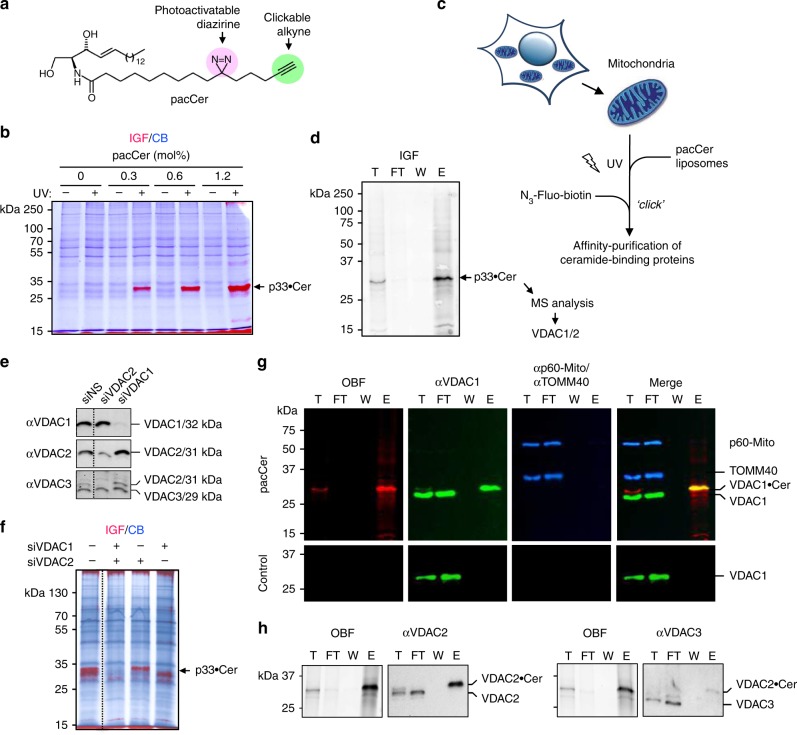
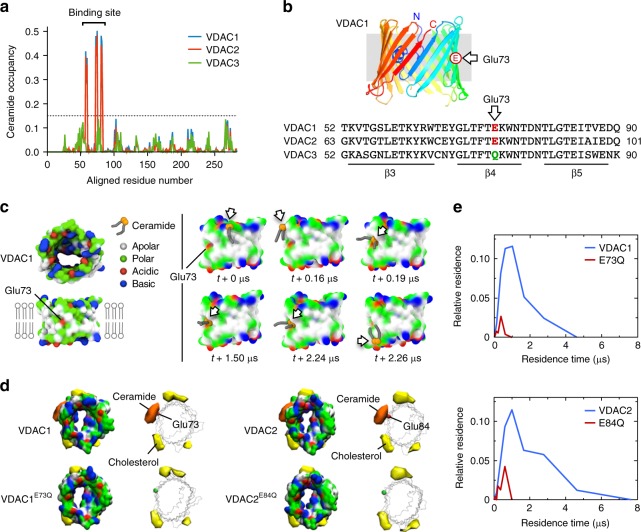
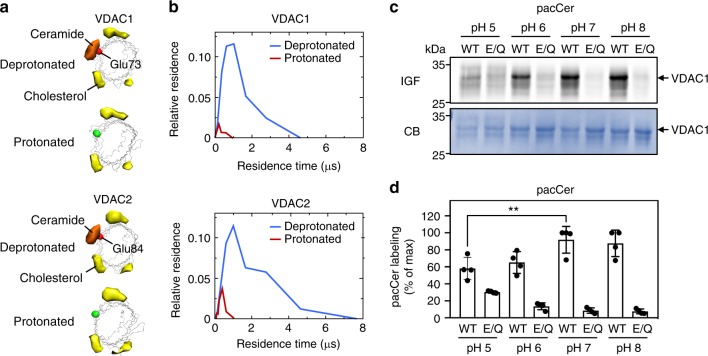
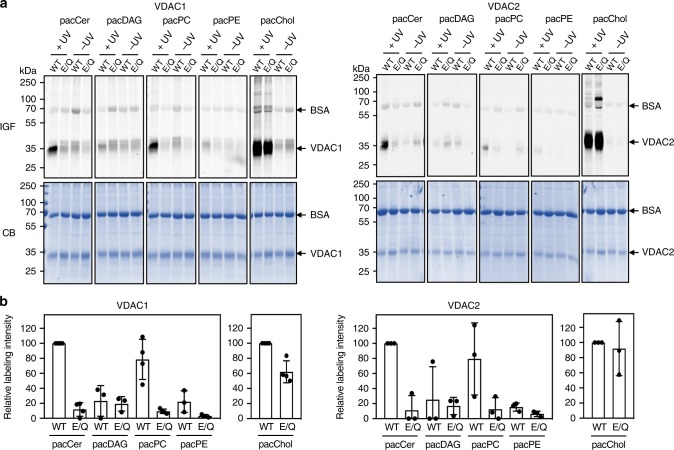
Comment in
-
Unraveling the molecular principles by which ceramides commit cells to death.Cell Stress. 2019 Jul 16;3(8):280-283. doi: 10.15698/cst2019.08.196. Cell Stress. 2019. PMID: 31440742 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
