

The Na+/H+-Exchanger NHE1 Regulates Extra- and Intracellular pH and Nimodipine-sensitive [Ca2+]i in the Suprachiasmatic Nucleus
- PMID: 31015514
- PMCID: PMC6478949
- DOI: 10.1038/s41598-019-42872-w
The Na+/H+-Exchanger NHE1 Regulates Extra- and Intracellular pH and Nimodipine-sensitive [Ca2+]i in the Suprachiasmatic Nucleus
Abstract
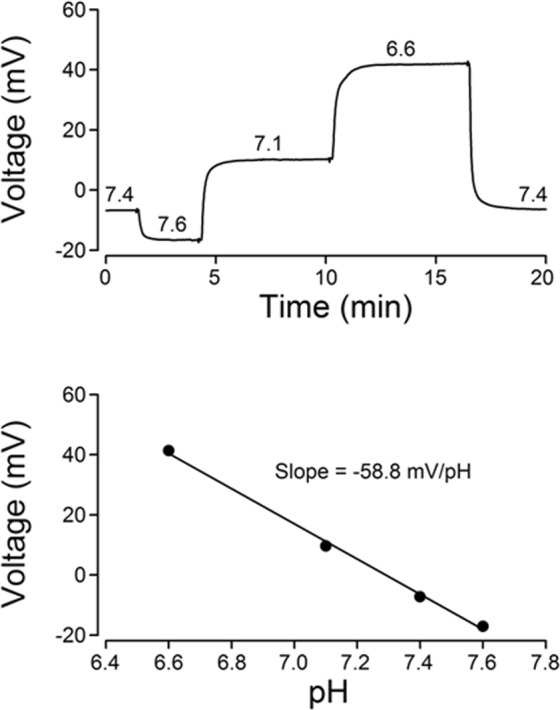
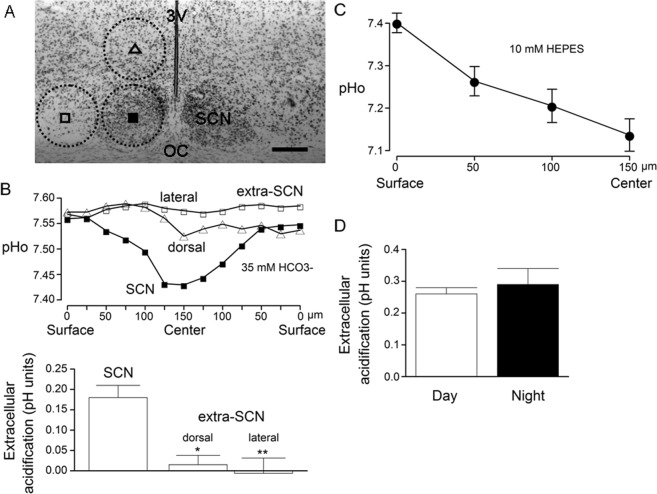
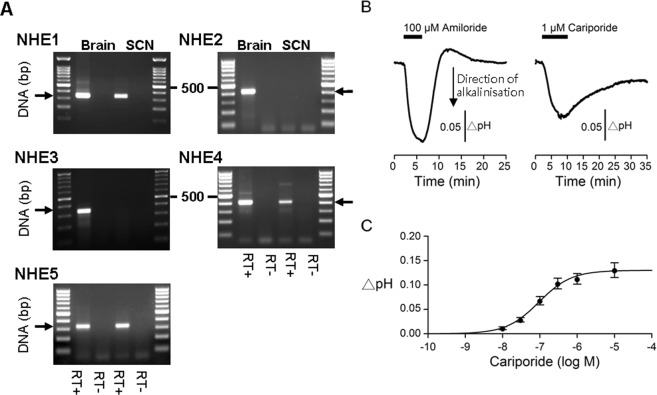
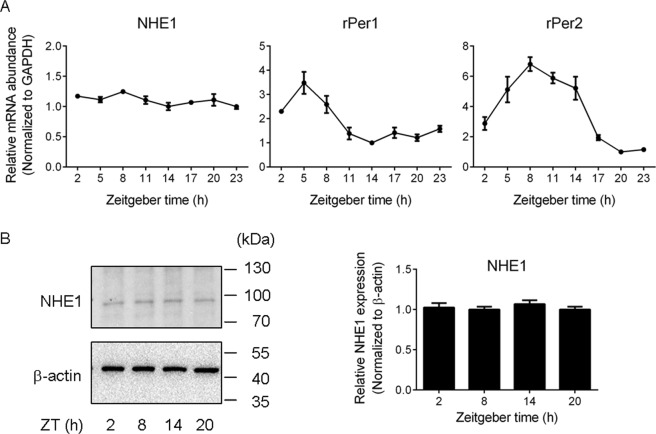
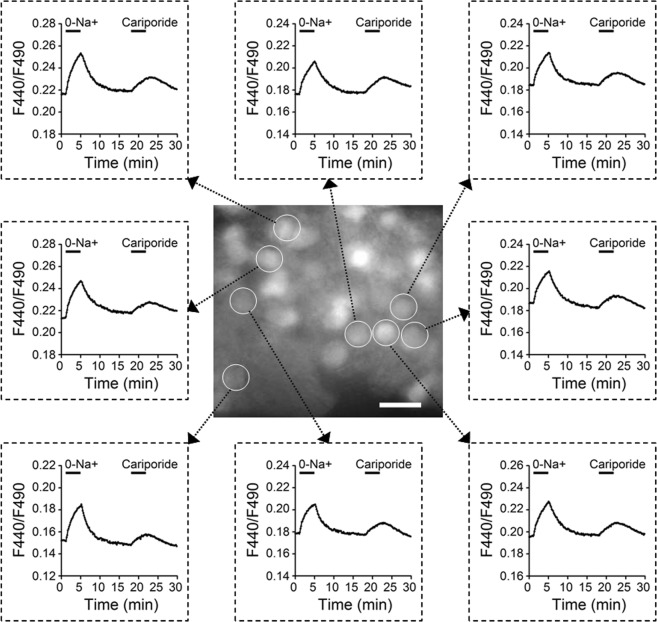
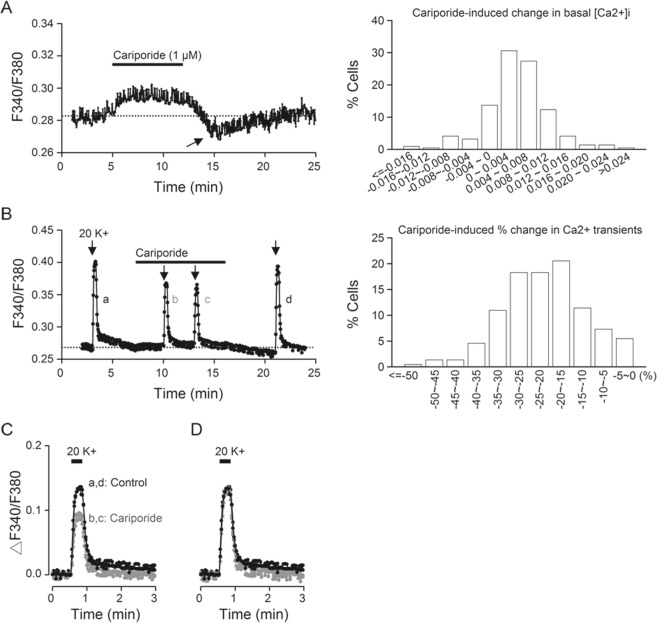
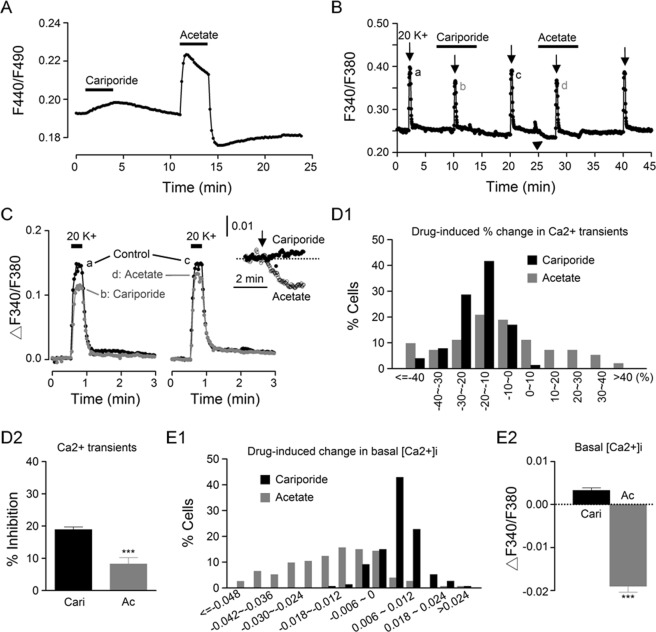
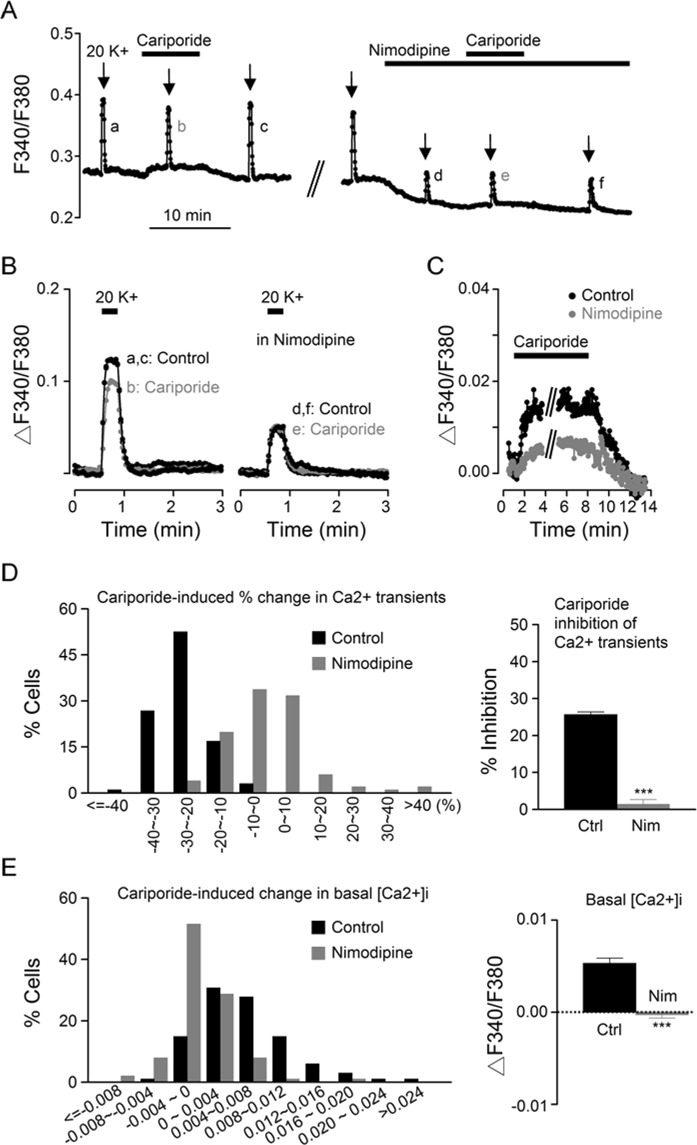
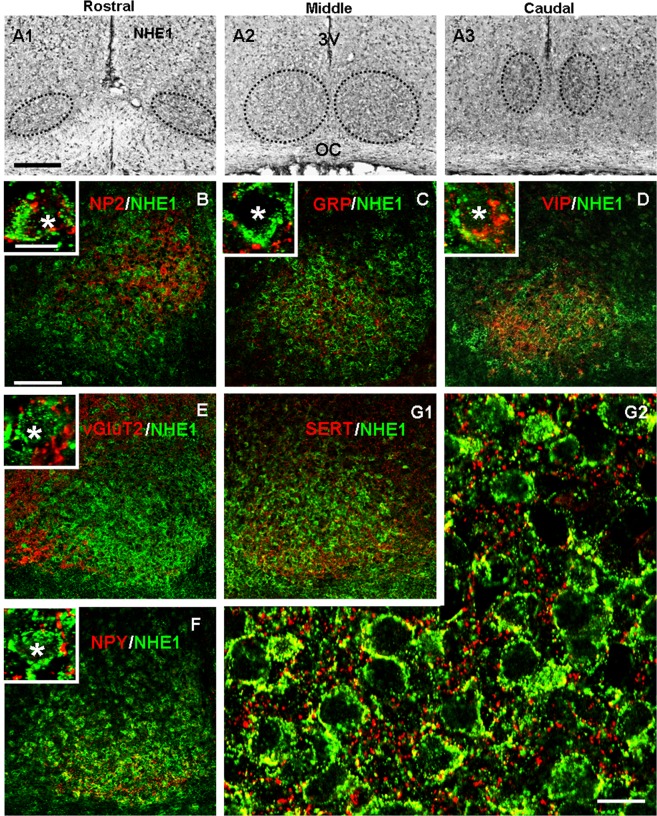
The central clock in the suprachiasmatic nucleus (SCN) has higher metabolic activity than extra-SCN areas in the anterior hypothalamus. Here we investigated whether the Na+/H+ exchanger (NHE) may regulate extracellular pH (pHe), intracellular pH (pHi) and [Ca2+]i in the SCN. In hypothalamic slices bathed in HEPES-buffered solution a standing acidification of ~0.3 pH units was recorded with pH-sensitive microelectrodes in the SCN but not extra-SCN areas. The NHE blocker amiloride alkalinised the pHe. RT-PCR revealed mRNA for plasmalemmal-type NHE1, NHE4, and NHE5 isoforms, whereas the NHE1-specific antagonist cariporide alkalinised the pHe. Real-time PCR and western blotting failed to detect day-night variation in NHE1 mRNA and protein levels. Cariporide induced intracellular acidosis, increased basal [Ca2+]i, and decreased depolarisation-induced Ca2+ rise, with the latter two effects being abolished with nimodipine blocking the L-type Ca2+ channels. Immunofluorescent staining revealed high levels of punctate colocalisation of NHE1 with serotonin transporter (SERT) or CaV1.2, as well as triple staining of NHE1, CaV1.2, and SERT or the presynaptic marker Bassoon. Our results indicate that NHE1 actively extrudes H+ to regulate pHi and nimodipine-sensitive [Ca2+]i in the soma, and along with CaV1.2 may also regulate presynaptic Ca2+ levels and, perhaps at least serotonergic, neurotransmission in the SCN.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
