

Acyloxyacyl hydrolase modulates depressive-like behaviors through aryl hydrocarbon receptor
- PMID: 31017816
- PMCID: PMC6732428
- DOI: 10.1152/ajpregu.00029.2019
Acyloxyacyl hydrolase modulates depressive-like behaviors through aryl hydrocarbon receptor
Abstract
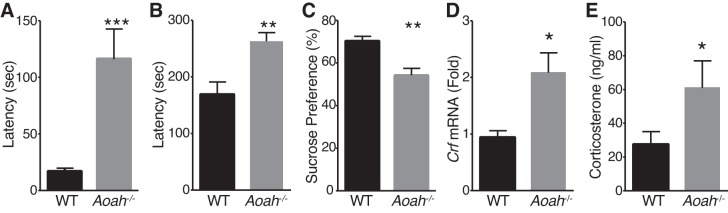
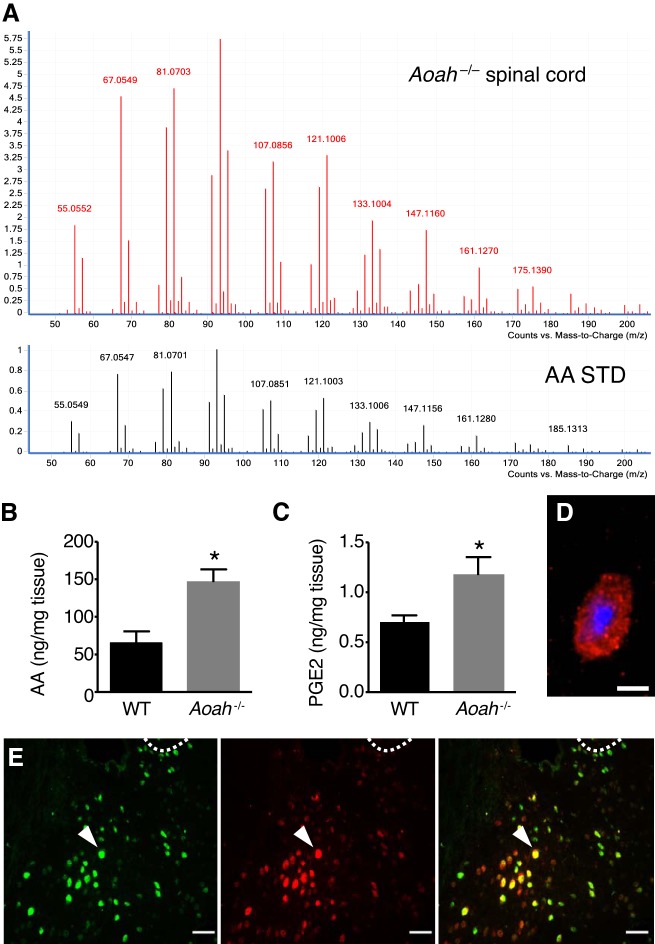
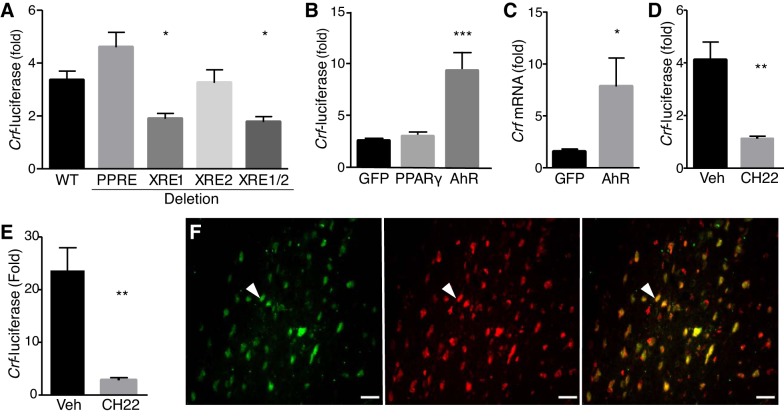
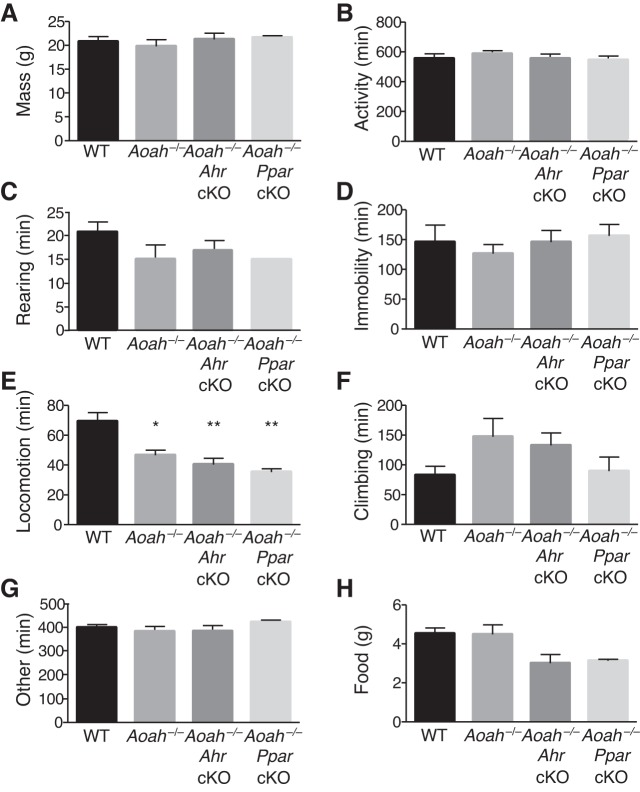
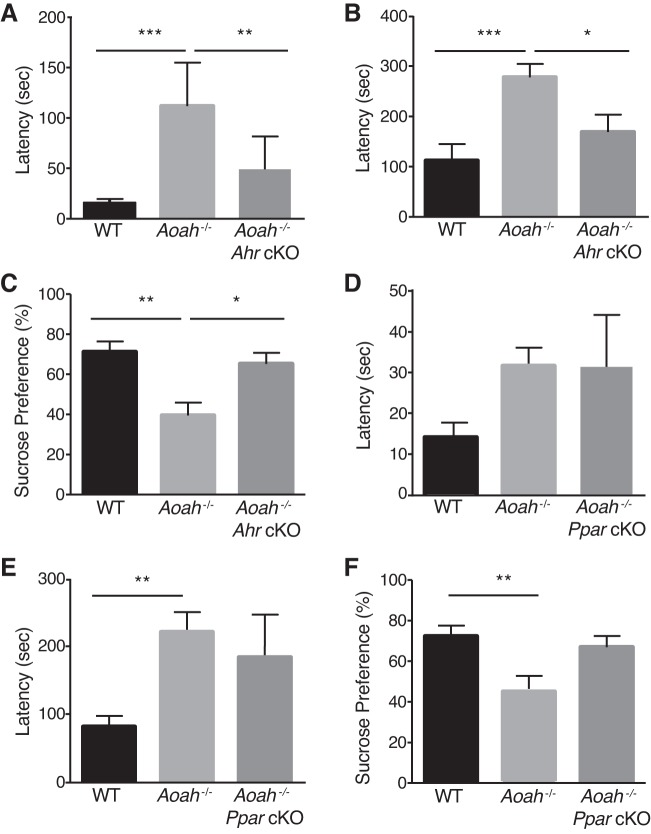
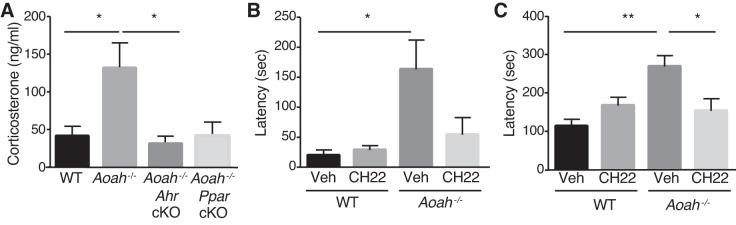
Corticotropin-releasing factor (CRF) regulates stress responses, and aberrant CRF signals are associated with depressive disorders. Crf expression is responsive to arachidonic acid (AA), where CRF is released from the hypothalamic paraventricular nucleus (PVN) to initiate the hypothalamic-pituitary-adrenal axis, culminating in glucocorticoid stress hormone release. Despite this biological and clinical significance, Crf regulation is unclear. Here, we report that acyloxyacyl hydrolase, encoded by Aoah, is expressed in the PVN, and Aoah regulates Crf through the aryl hydrocarbon receptor (AhR). We previously showed that AOAH-deficient mice mimicked interstitial cystitis/bladder pain syndrome, a condition frequently associated with comorbid anxiety and depression. With the use of novelty-suppressed feeding and sucrose preference assays to quantify rodent correlates of anxiety/depression, AOAH-deficient mice exhibited depressive behaviors. AOAH-deficient mice also had increased CNS AA, increased Crf expression in the PVN, and elevated serum corticosterone, consistent with dysfunction of the hypothalamic-pituitary-adrenal axis. The human Crf promoter has putative binding sites for AhR and peroxisome proliferator-activated receptor (PPARγ). PPARγ did not affect AA-dependent Crf expression in vitro, and conditional Pparγ knockout did not alter the AOAH-deficient depressive phenotype, despite previous studies implicating PPARγ as a therapeutic target for depression. In contrast, Crf induction was mediated by AhR binding sites in vitro and increased by AhR overexpression. Furthermore, conditional Ahr knockout rescued the depressive phenotype of AOAH-deficient mice. Finally, an AhR antagonist rescued the AOAH-deficient depressive phenotype. Together, our results demonstrate that Aoah is a novel genetic regulator of Crf mediated through AhR, and AhR is a therapeutic target for depression.
Keywords: AOAH; AhR; CRF; arachidonic acid; interstitial cystitis; stress.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures

References
-
- Agudelo LZ, Femenía T, Orhan F, Porsmyr-Palmertz M, Goiny M, Martinez-Redondo V, Correia JC, Izadi M, Bhat M, Schuppe-Koistinen I, Pettersson AT, Ferreira DMS, Krook A, Barres R, Zierath JR, Erhardt S, Lindskog M, Ruas JL. Skeletal muscle PGC-1α1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 159: 33–45, 2014. [Erratum in Cell 160: P351, 2015]. doi: 10.1016/j.cell.2014.07.051. - DOI - PubMed
-
- Alesci S, Martinez PE, Kelkar S, Ilias I, Ronsaville DS, Listwak SJ, Ayala AR, Licinio J, Gold HK, Kling MA, Chrousos GP, Gold PW. Major depression is associated with significant diurnal elevations in plasma interleukin-6 levels, a shift of its circadian rhythm, and loss of physiological complexity in its secretion: clinical implications. J Clin Endocrinol Metab 90: 2522–2530, 2005. doi: 10.1210/jc.2004-1667. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases