

CD34 defines melanocyte stem cell subpopulations with distinct regenerative properties
- PMID: 31017901
- PMCID: PMC6481766
- DOI: 10.1371/journal.pgen.1008034
CD34 defines melanocyte stem cell subpopulations with distinct regenerative properties
Abstract
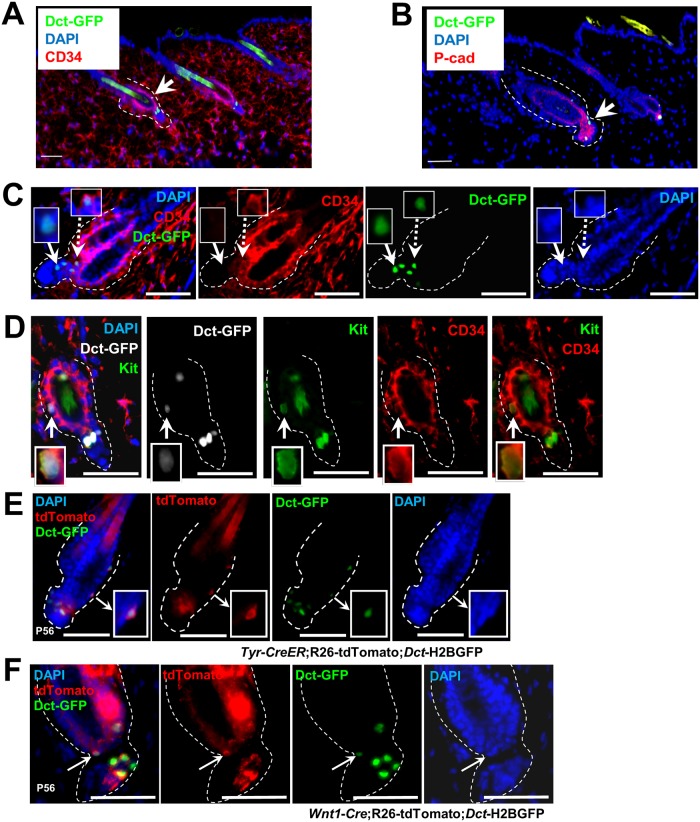
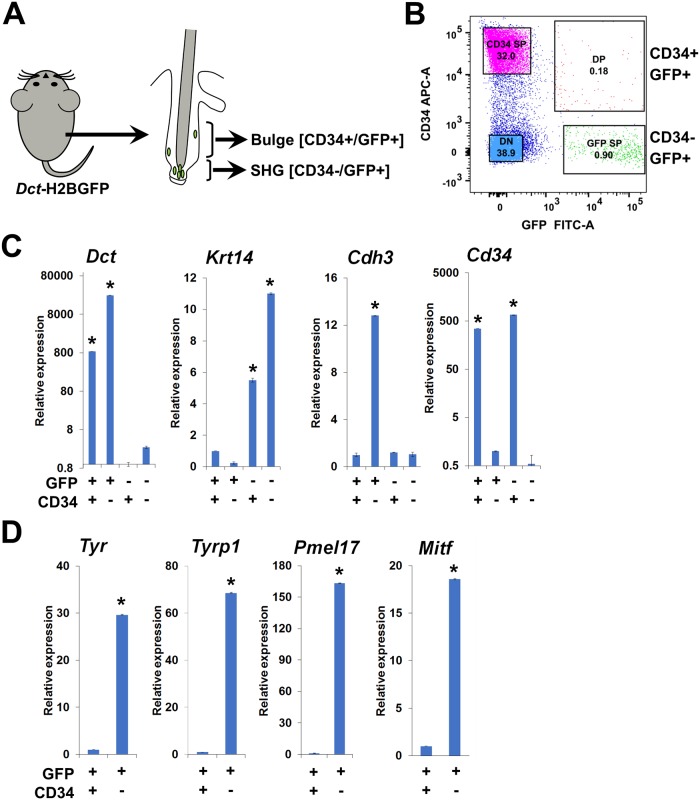
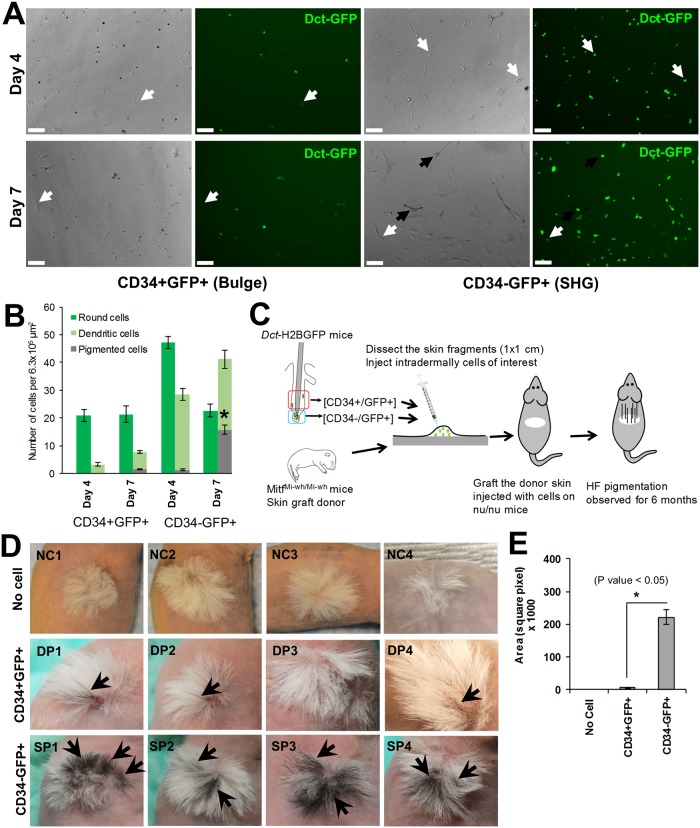
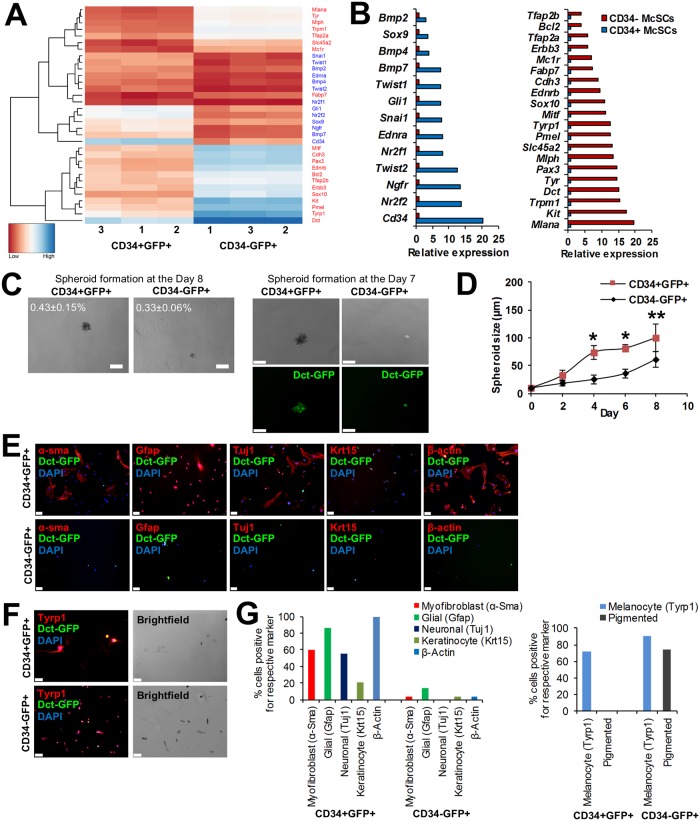
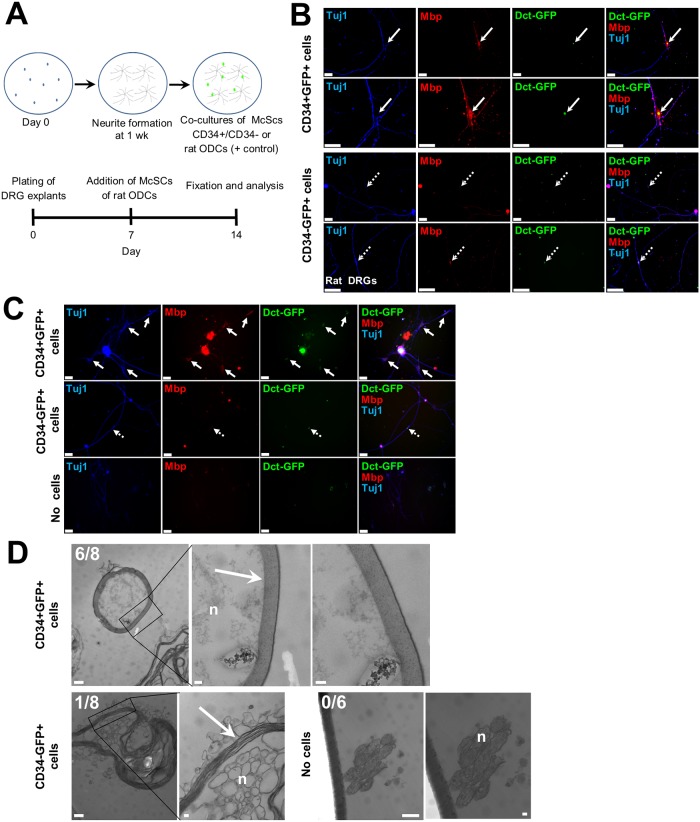
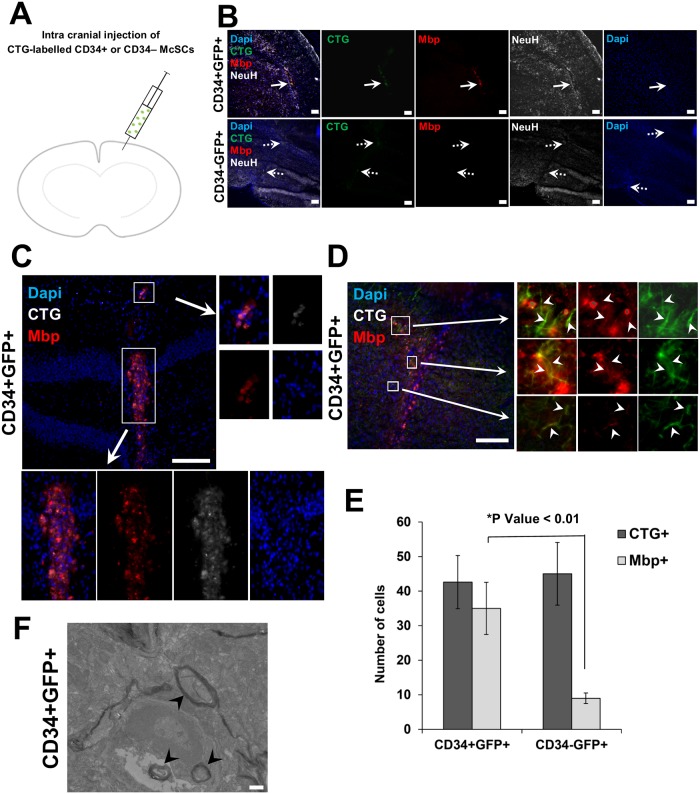
Melanocyte stem cells (McSCs) are the undifferentiated melanocytic cells of the mammalian hair follicle (HF) responsible for recurrent generation of a large number of differentiated melanocytes during each HF cycle. HF McSCs reside in both the CD34+ bulge/lower permanent portion (LPP) and the CD34- secondary hair germ (SHG) regions of the HF during telogen. Using Dct-H2BGFP mice, we separate bulge/LPP and SHG McSCs using FACS with GFP and anti-CD34 to show that these two subsets of McSCs are functionally distinct. Genome-wide expression profiling results support the distinct nature of these populations, with CD34- McSCs exhibiting higher expression of melanocyte differentiation genes and with CD34+ McSCs demonstrating a profile more consistent with a neural crest stem cell. In culture and in vivo, CD34- McSCs regenerate pigmentation more efficiently whereas CD34+ McSCs selectively exhibit the ability to myelinate neurons. CD34+ McSCs, and their counterparts in human skin, may be useful for myelinating neurons in vivo, leading to new therapeutic opportunities for demyelinating diseases and traumatic nerve injury.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
