

Disruption in A-to-I Editing Levels Affects C. elegans Development More Than a Complete Lack of Editing
- PMID: 31018137
- PMCID: PMC8139731
- DOI: 10.1016/j.celrep.2019.03.095
Disruption in A-to-I Editing Levels Affects C. elegans Development More Than a Complete Lack of Editing
Abstract
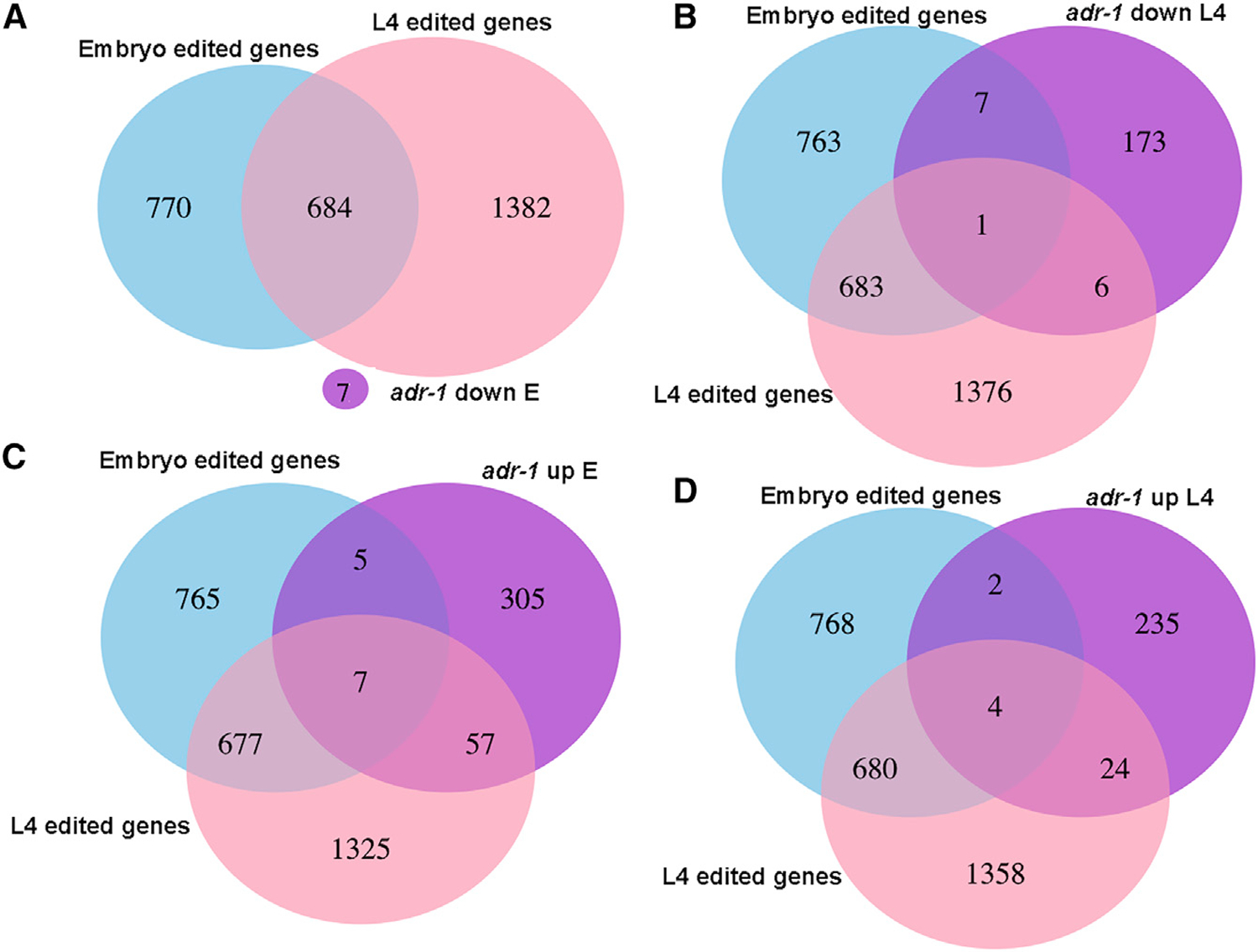
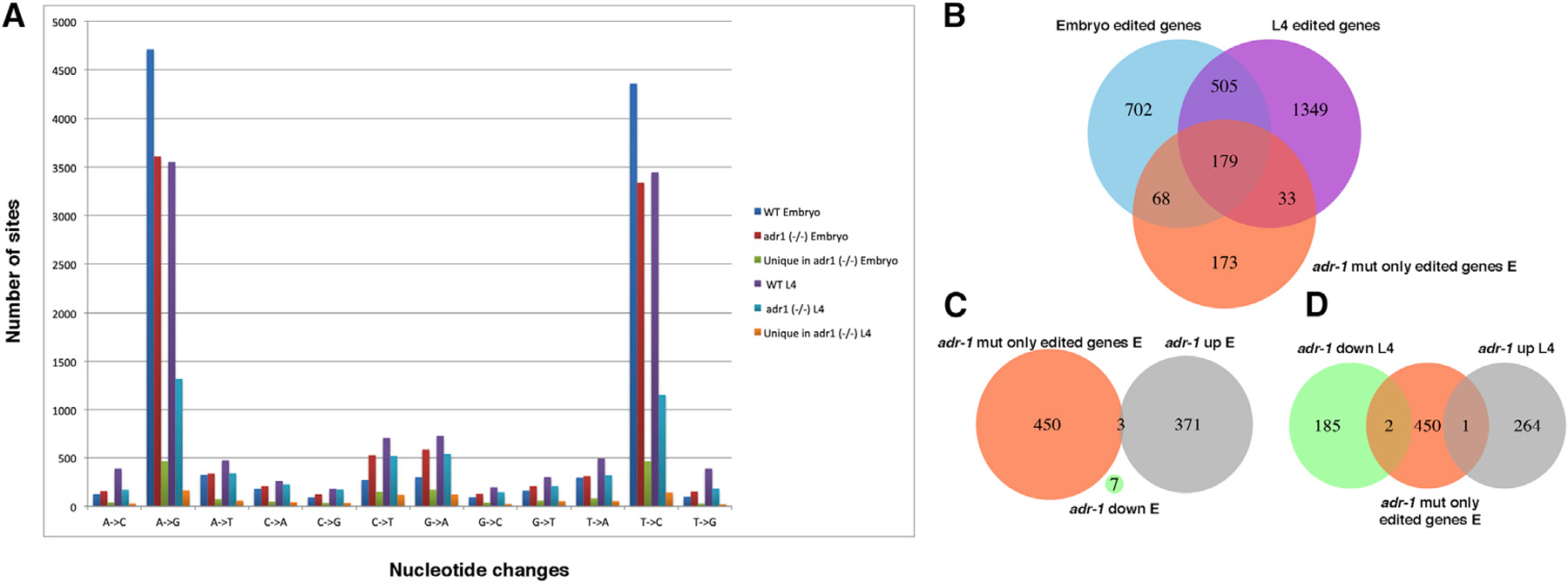
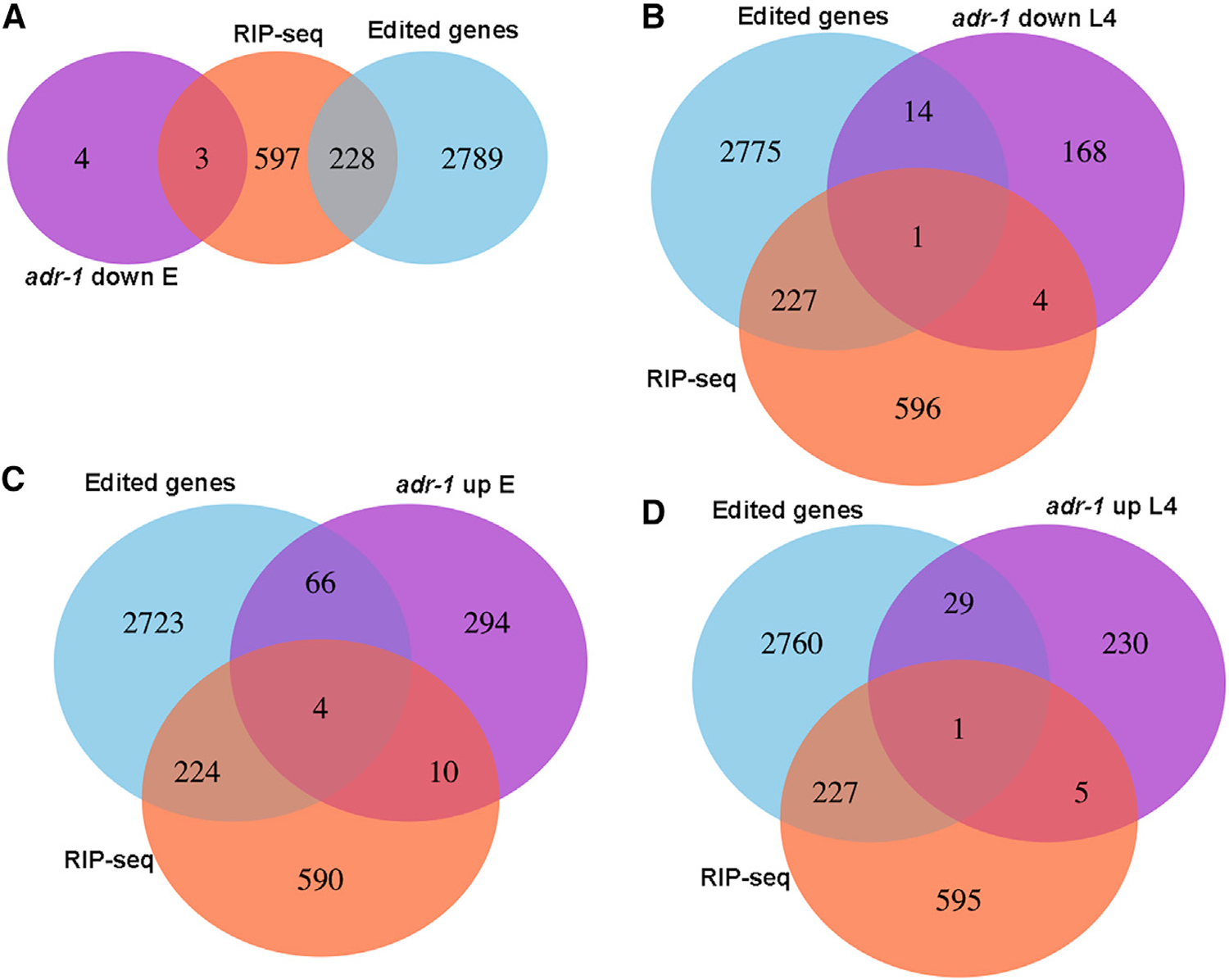
A-to-I RNA editing, catalyzed by ADAR proteins, is widespread in eukaryotic transcriptomes. Studies showed that, in C. elegans, ADR-2 can actively deaminate dsRNA, whereas ADR-1 cannot. Therefore, we set out to study the effect of each of the ADAR genes on the RNA editing process. We performed comprehensive phenotypic, transcriptomics, proteomics, and RNA binding screens on worms mutated in a single ADAR gene. We found that ADR-1 mutants exhibit more-severe phenotypes than ADR-2, and some of them are a result of non-editing functions of ADR-1. We also show that ADR-1 significantly binds edited genes and regulates mRNA expression, whereas the effect on protein levels is minor. In addition, ADR-1 primarily promotes editing by ADR-2 at the L4 stage of development. Our results suggest that ADR-1 has a significant role in the RNA editing process and in altering editing levels that affect RNA expression; loss of ADR-1 results in severe phenotypes.
Keywords: ADAR; C. elegans; gene expression; organism development; transcriptomics.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
ADBP-1 regulates ADR-2 nuclear localization to control editing substrate selection.Nucleic Acids Res. 2024 Sep 9;52(16):9501-9518. doi: 10.1093/nar/gkae641. Nucleic Acids Res. 2024. PMID: 39036970 Free PMC article.
-
The dsRBP and inactive editor ADR-1 utilizes dsRNA binding to regulate A-to-I RNA editing across the C. elegans transcriptome.Cell Rep. 2014 Feb 27;6(4):599-607. doi: 10.1016/j.celrep.2014.01.011. Epub 2014 Feb 6. Cell Rep. 2014. PMID: 24508457 Free PMC article.
-
Profiling the RNA editomes of wild-type C. elegans and ADAR mutants.Genome Res. 2015 Jan;25(1):66-75. doi: 10.1101/gr.176107.114. Epub 2014 Nov 4. Genome Res. 2015. PMID: 25373143 Free PMC article.
-
A-to-I RNA editing - thinking beyond the single nucleotide.RNA Biol. 2017 Dec 2;14(12):1690-1694. doi: 10.1080/15476286.2017.1364830. Epub 2017 Oct 11. RNA Biol. 2017. PMID: 28820319 Free PMC article. Review.
-
Bioinformatic approaches for accurate assessment of A-to-I editing in complete transcriptomes.Methods Enzymol. 2025;710:241-265. doi: 10.1016/bs.mie.2024.11.020. Epub 2024 Nov 23. Methods Enzymol. 2025. PMID: 39870448 Review.
Cited by
-
Lactobacillus for ribosome peptide editing cancer.Clin Transl Oncol. 2023 Jun;25(6):1522-1544. doi: 10.1007/s12094-022-03066-5. Epub 2023 Jan 24. Clin Transl Oncol. 2023. PMID: 36694080 Free PMC article. Review.
-
Caenorhabditis elegans inositol hexaphosphate pathways couple to RNA interference and pathogen defense.Proc Natl Acad Sci U S A. 2024 Dec 3;121(49):e2416982121. doi: 10.1073/pnas.2416982121. Epub 2024 Nov 27. Proc Natl Acad Sci U S A. 2024. PMID: 39602251 Free PMC article.
-
Full-length direct RNA sequencing reveals extensive remodeling of RNA expression, processing and modification in aging Caenorhabditis elegans.Nucleic Acids Res. 2024 Dec 11;52(22):13896-13913. doi: 10.1093/nar/gkae1064. Nucleic Acids Res. 2024. PMID: 39558169 Free PMC article.
-
ADBP-1 regulates ADR-2 nuclear localization to control editing substrate selection.Nucleic Acids Res. 2024 Sep 9;52(16):9501-9518. doi: 10.1093/nar/gkae641. Nucleic Acids Res. 2024. PMID: 39036970 Free PMC article.
-
ADAR-mediated regulation of PQM-1 expression in neurons impacts gene expression throughout C. elegans and regulates survival from hypoxia.PLoS Biol. 2023 Sep 25;21(9):e3002150. doi: 10.1371/journal.pbio.3002150. eCollection 2023 Sep. PLoS Biol. 2023. PMID: 37747897 Free PMC article.
References
-
- Benian GM, Kiff JE, Neckelmann N, Moerman DG, and Waterston RH (1989). Sequence of an unusually large protein implicated in regulation of myosin activity in C. elegans. Nature 342, 45–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
