

The Major Risk Factors for Alzheimer's Disease: Age, Sex, and Genes Modulate the Microglia Response to Aβ Plaques
- PMID: 31018141
- PMCID: PMC7340153
- DOI: 10.1016/j.celrep.2019.03.099
The Major Risk Factors for Alzheimer's Disease: Age, Sex, and Genes Modulate the Microglia Response to Aβ Plaques
Abstract
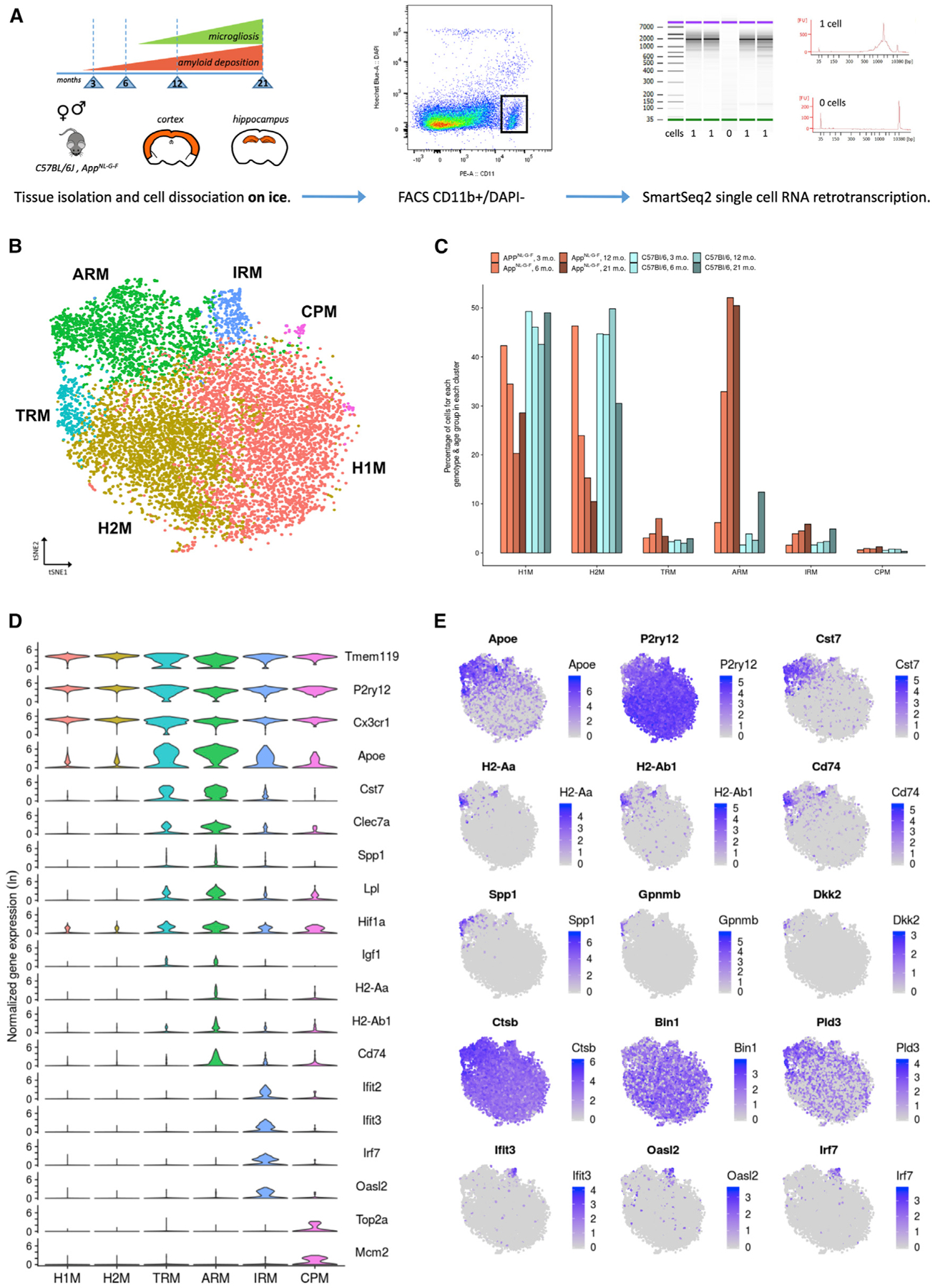
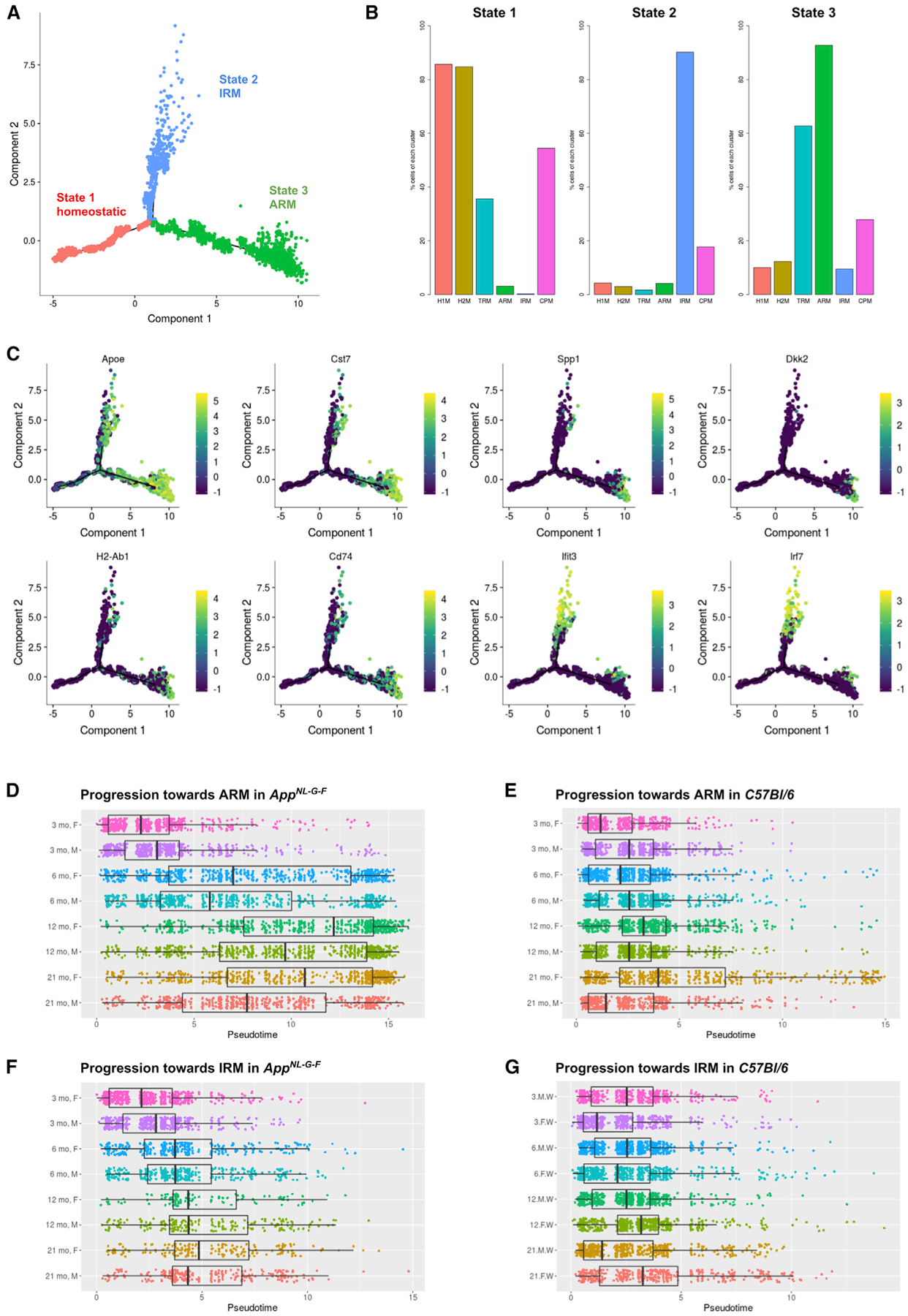
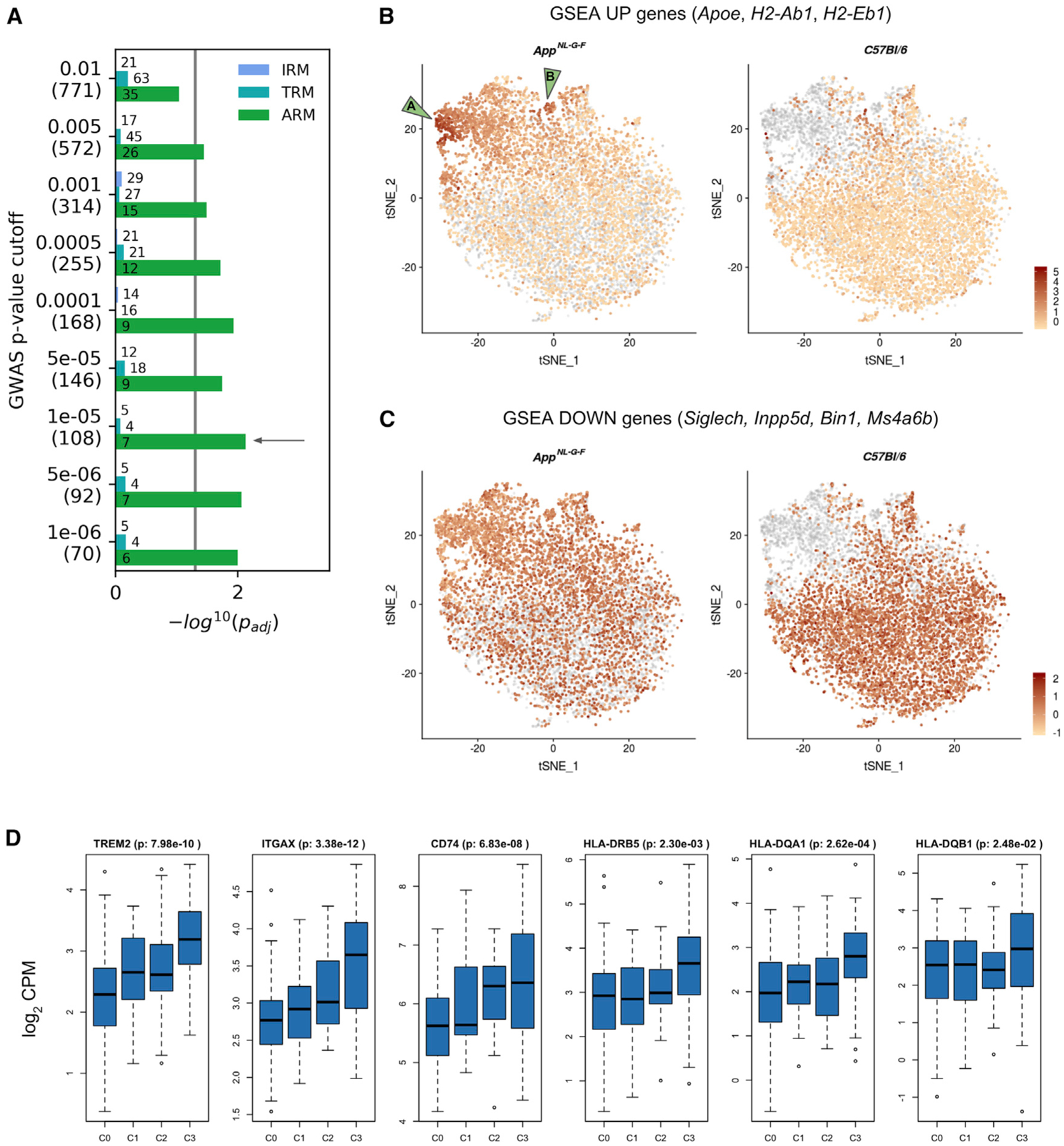
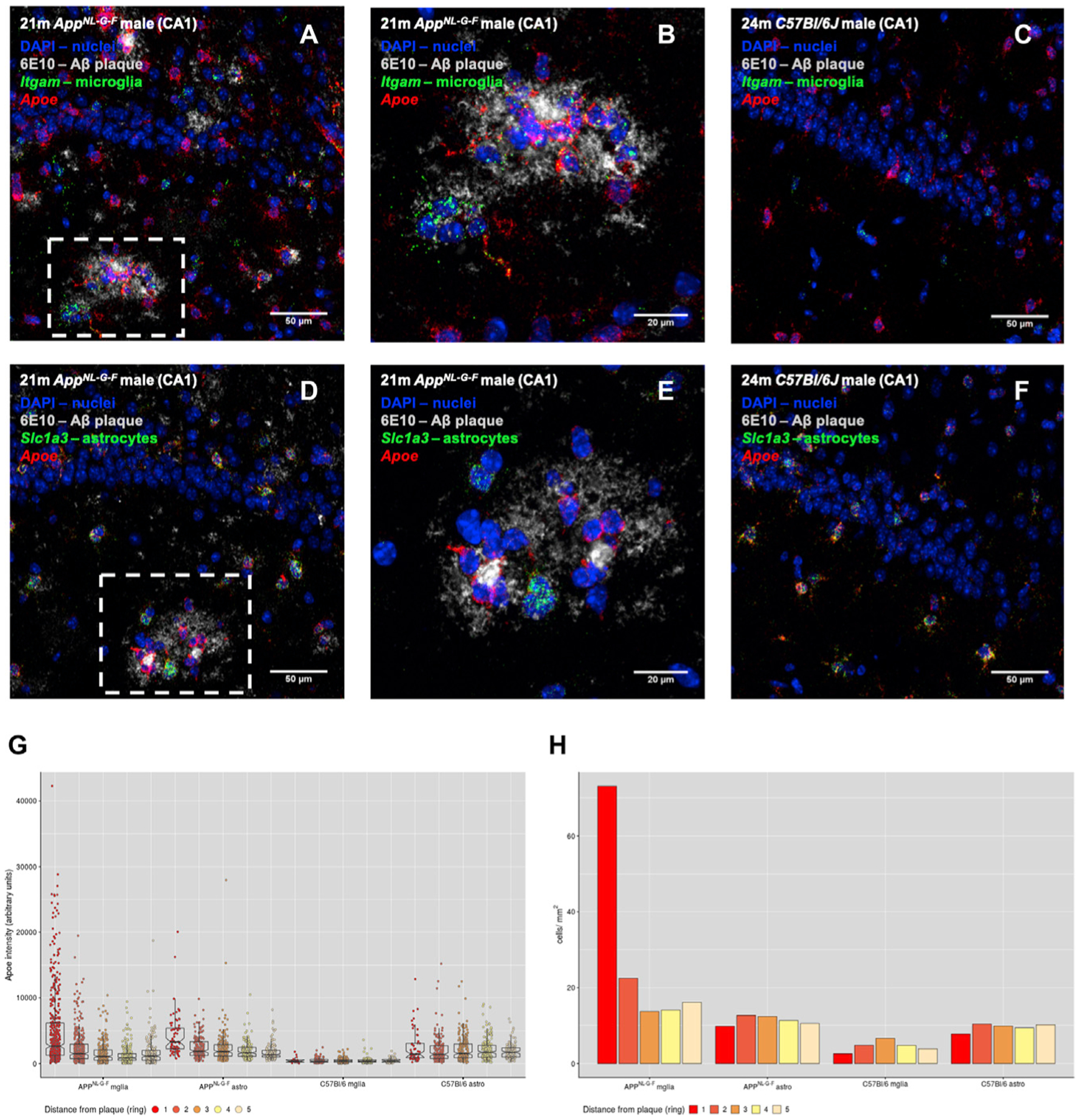
Gene expression profiles of more than 10,000 individual microglial cells isolated from cortex and hippocampus of male and female AppNL-G-F mice over time demonstrate that progressive amyloid-β accumulation accelerates two main activated microglia states that are also present during normal aging. Activated response microglia (ARMs) are composed of specialized subgroups overexpressing MHC type II and putative tissue repair genes (Dkk2, Gpnmb, and Spp1) and are strongly enriched with Alzheimer's disease (AD) risk genes. Microglia from female mice progress faster in this activation trajectory. Similar activated states are also found in a second AD model and in human brain. Apoe, the major genetic risk factor for AD, regulates the ARMs but not the interferon response microglia (IRMs). Thus, the ARMs response is the converging point for aging, sex, and genetic AD risk factors.
Keywords: ARM; Alzheimer; IRM; apoe; app knock in; in situ RNA hybridization; microglia; single cell RNA-seq; single cell sequencing.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
M.E.W., G.S., T.M., and E.K. are employed by AbbVie, Inc. AbbVie, Inc., has subsidized part of the study. B.D.S. is a consultant for several companies. All the other authors declare no competing interests.
Figures
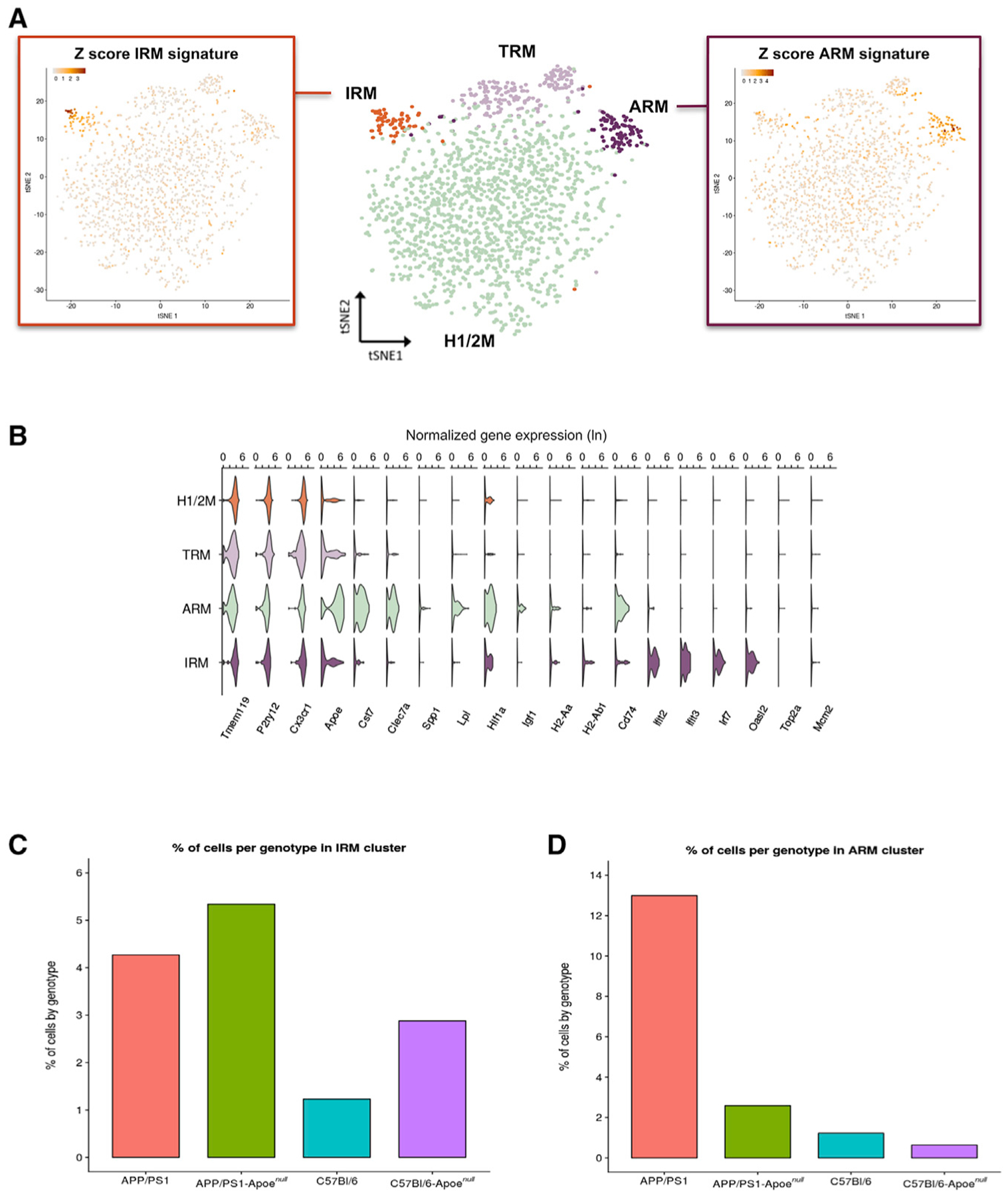
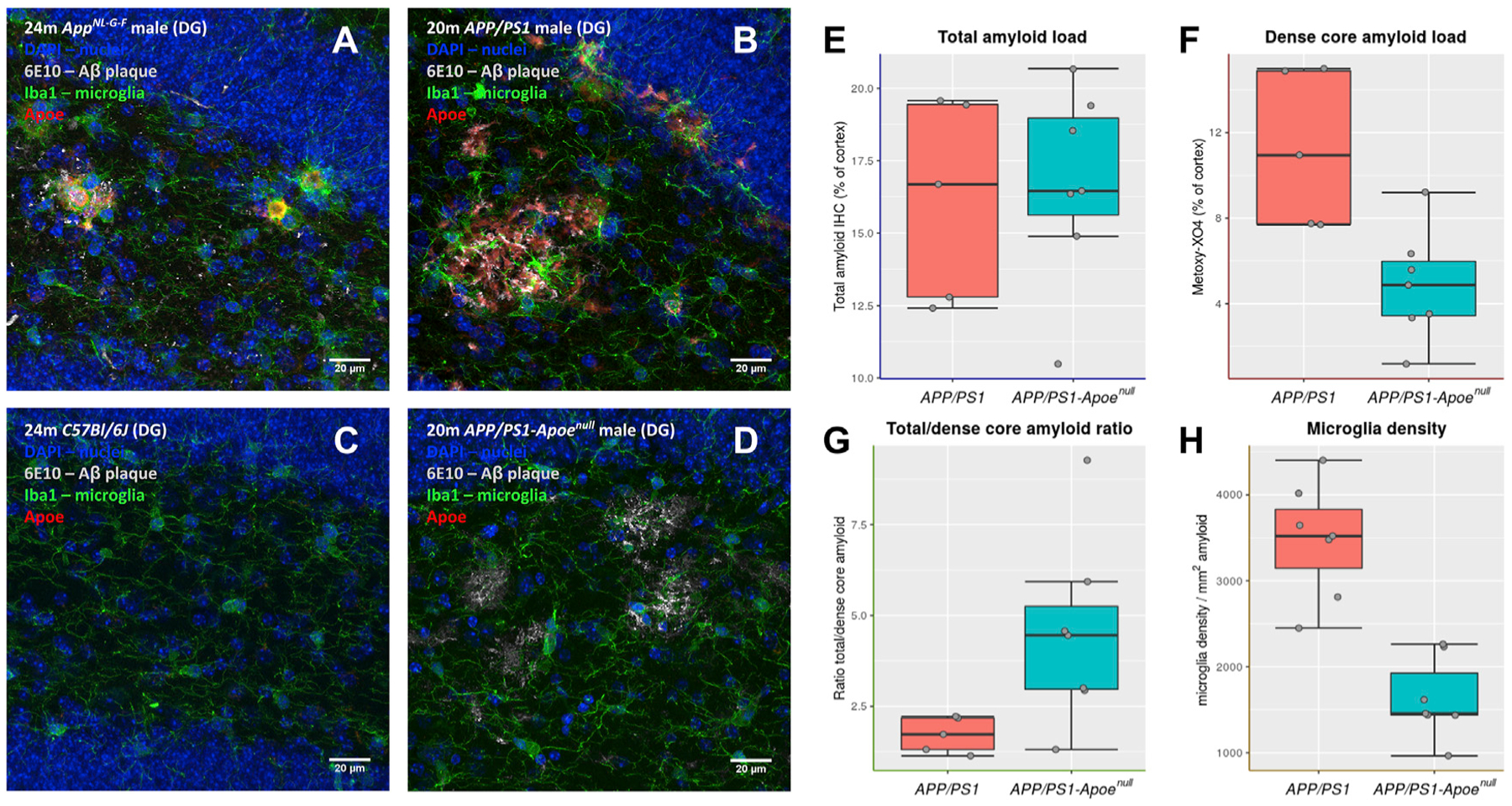
Comment in
-
Age-related microglial activation accelerated in AD.Nat Rev Neurol. 2019 Jul;15(7):369. doi: 10.1038/s41582-019-0204-2. Nat Rev Neurol. 2019. PMID: 31086265 No abstract available.
References
-
- Behl C, and Ziegler C (2017). Beyond amyloid—widening the view on Alzheimer’s disease. J. Neurochem 143, 394–395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
