

Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species
- PMID: 31018580
- PMCID: PMC6523773
- DOI: 10.3390/biom9040159
Curcuma raktakanda Induces Apoptosis and Suppresses Migration in Cancer Cells: Role of Reactive Oxygen Species
Abstract
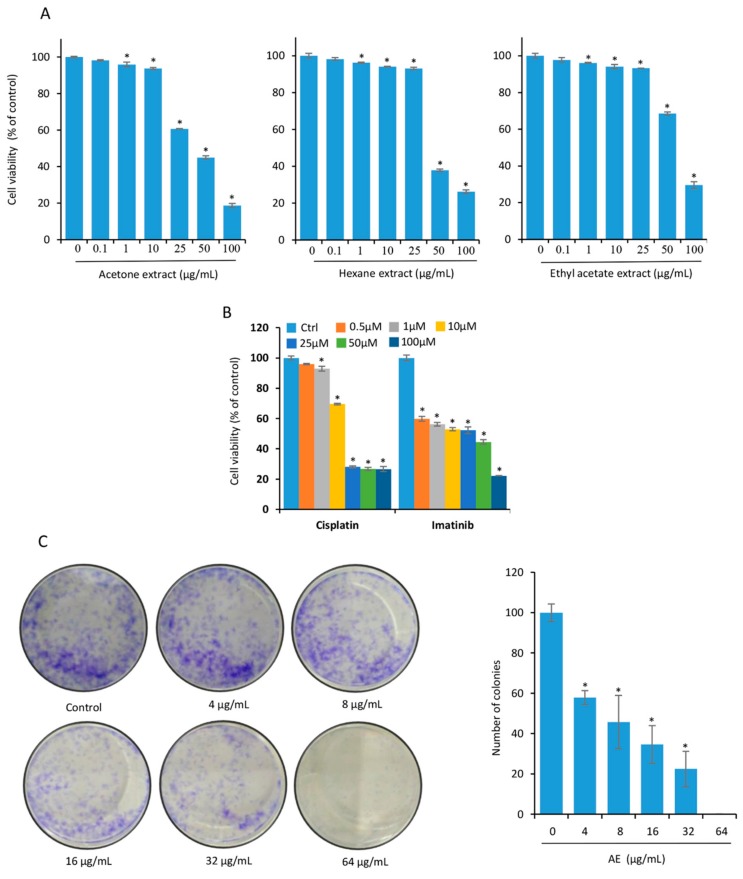
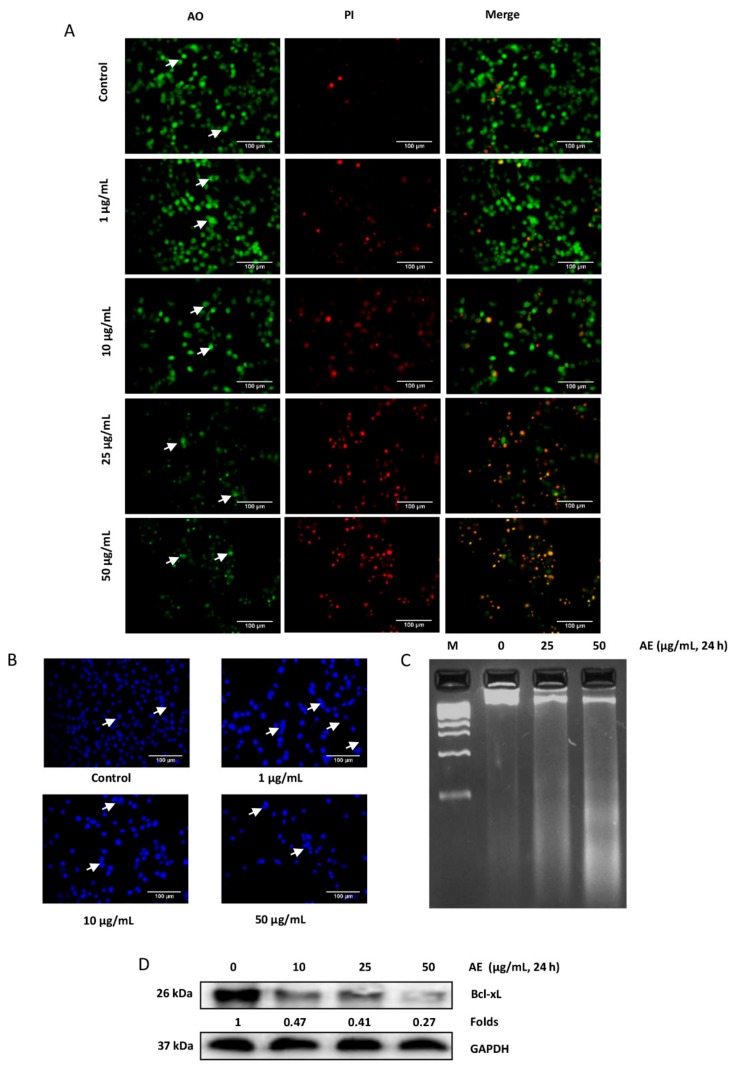
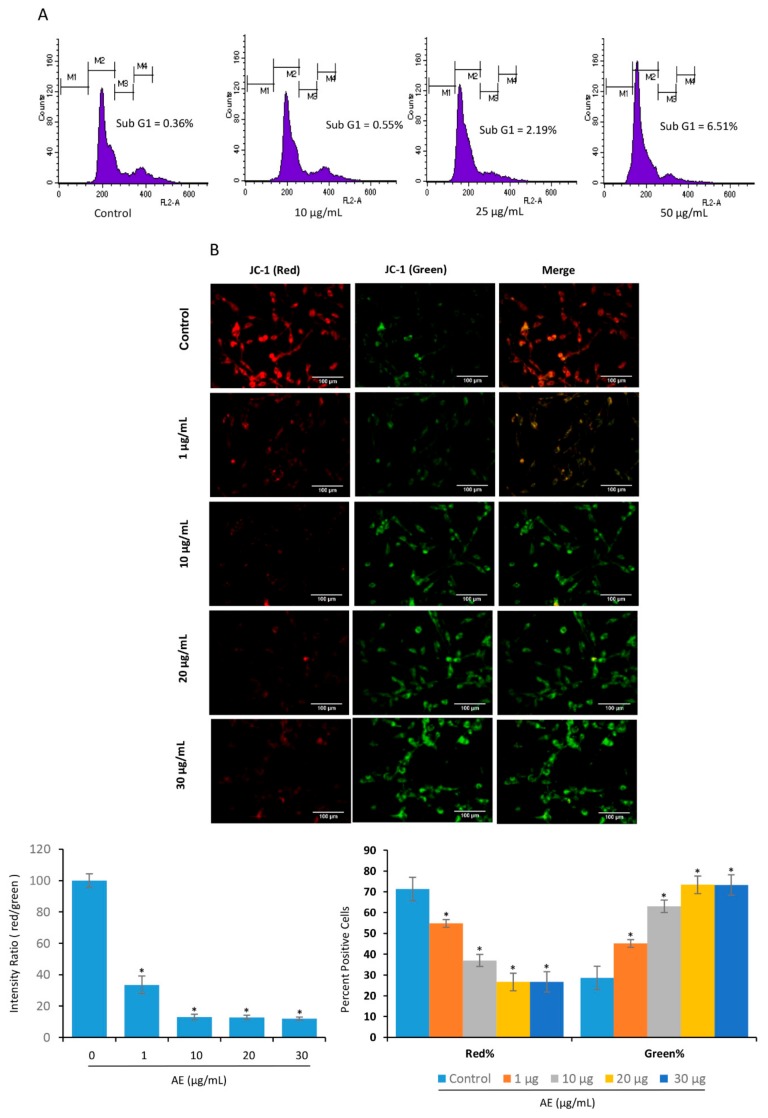
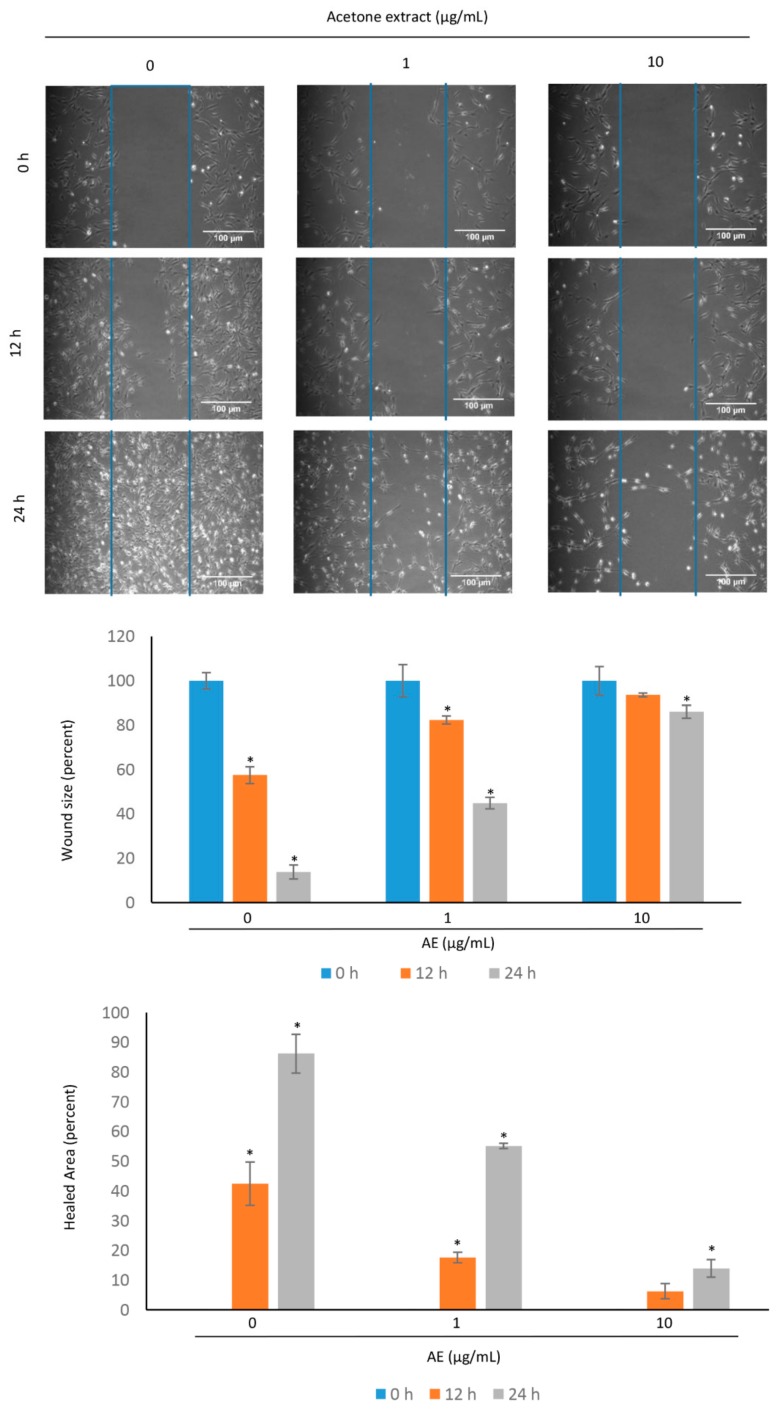
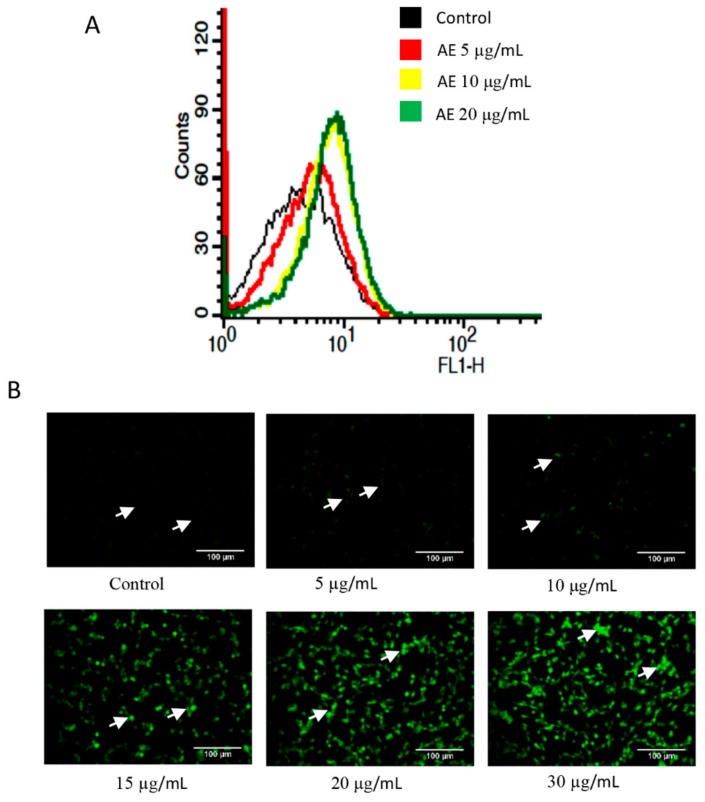
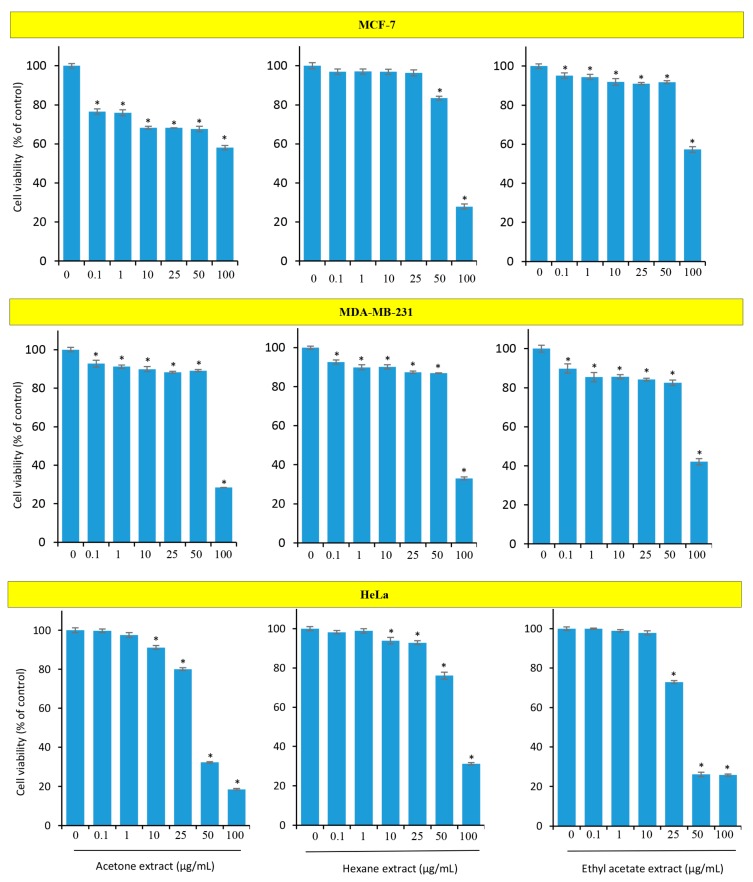
Although over 100 species of Curcuma are reported, only Curcuma longa is extensively studied. Curcuma raktakanda, a poorly studied species, is most commonly distributed in the Kerala state of India. For the first time, we examined the efficacy of different fractions (acetone, hexane, and ethyl acetate) of C. raktakanda against glioma, cervical, and breast cancer cell lines. As determined by mitochondrial reductase activity assay, the viability of cancer cells was decreased in a concentration-dependent manner by the three fractions. The half maximal inhibitory concentration (IC-50) values after the treatment of C-6 glioma cells for 48 h was found to be 32.97 µg/mL (acetone extract), 40.63 µg/mL (hexane extract), and 51.65 µg/mL (ethyl acetate extract). Of the three fractions, the acetone fraction was more effective. The long-term colony formation of cancer cells was significantly suppressed by the acetone fraction. Analyses using DAPI (4',6-diamidino-2-phenylindole) staining, AO/PI (acridine orange/propidium iodide) staining, DNA laddering, and sub-G1 population revealed that the acetone extract induced apoptosis in glioma cells. The extract induced reactive oxygen species generation and suppressed the expression of cell survival proteins. The migration of cancer cells was also suppressed by the acetone extract. The gas chromatography-mass spectrometry (GC-MS) analysis indicated that tetracontane, dotriacontane, hexatriacontane, pentacosane, hexacosane, and eicosane are the major components in the acetone extract. Collectively, the extract from C. raktakanda exhibited anti-carcinogenic activities in cancer cells. We are exploring whether the phytoconstituents, individually, or collectively contribute to the anti-cancer activities of C. raktakanda.
Keywords: cancer; curcuma; glioblastoma; inflammation; reactive oxygen species.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Louis D.N., Perry A., Reifenberger G., Von Deimling A., Figarella-Branger D., Cavenee W.K., Ohgaki H., Wiestler O.D., Kleihues P., Ellison D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016;131:803–820. doi: 10.1007/s00401-016-1545-1. - DOI - PubMed
-
- Mirimanoff R.-O., Gorlia T., Mason W., Van den Bent M.J., Kortmann R.-D., Fisher B., Reni M., Brandes A.A., Curschmann J., Villa S. Radiotherapy and temozolomide for newly diagnosed glioblastoma: Recursive partitioning analysis of the EORTC 26981/22981-NCIC CE3 phase III randomized trial. J. Clin. Oncol. 2006;24:2563–2569. doi: 10.1200/JCO.2005.04.5963. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
