

Soluble Heparin and Heparan Sulfate Glycosaminoglycans Interfere with Sonic Hedgehog Solubilization and Receptor Binding
- PMID: 31018591
- PMCID: PMC6526471
- DOI: 10.3390/molecules24081607
Soluble Heparin and Heparan Sulfate Glycosaminoglycans Interfere with Sonic Hedgehog Solubilization and Receptor Binding
Abstract
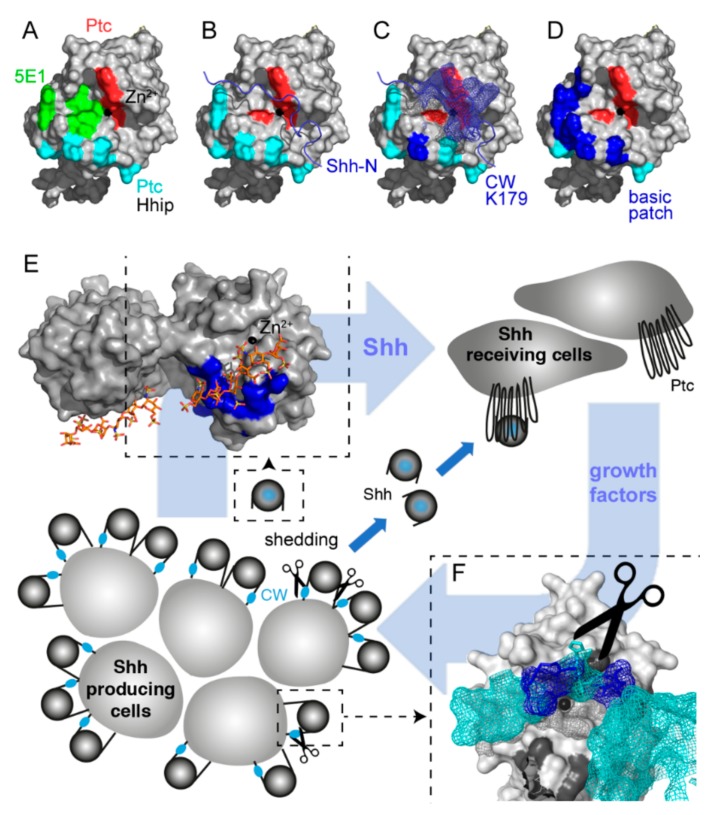
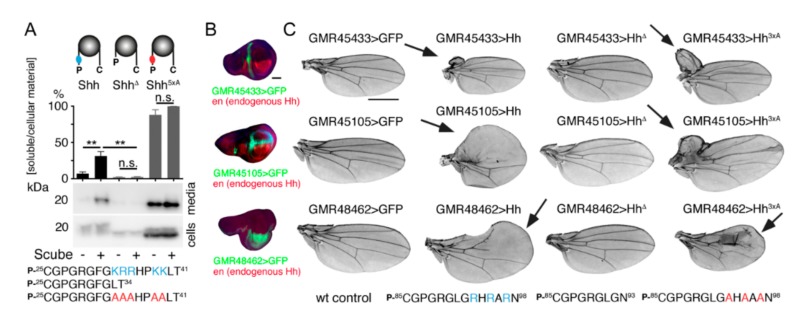
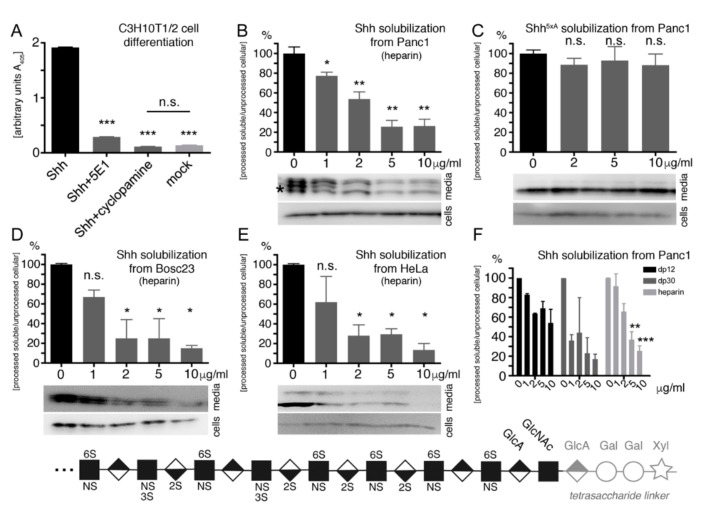
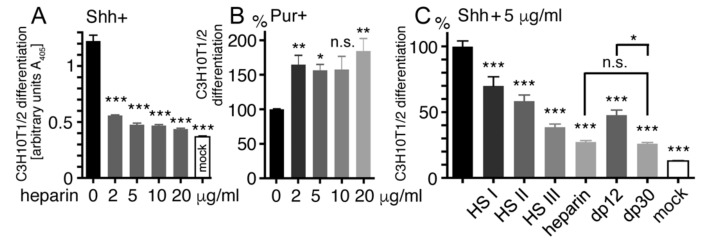
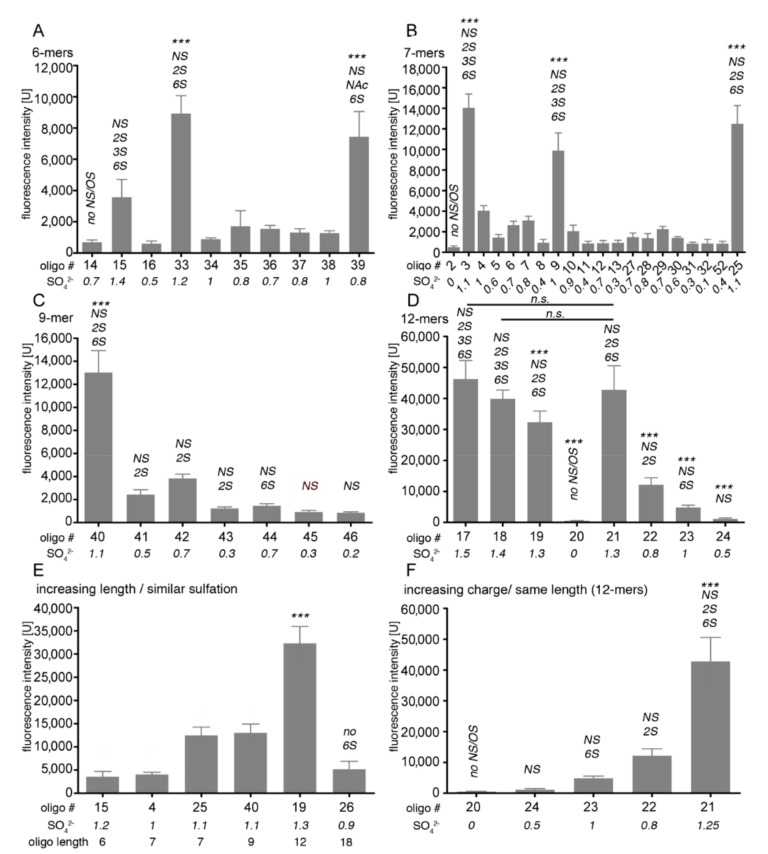
Sonic hedgehog (Shh) signaling plays a tumor-promoting role in many epithelial cancers. Cancer cells produce soluble a Shh that signals to distant stromal cells that express the receptor Patched (Ptc). These receiving cells respond by producing other soluble factors that promote cancer cell growth, generating a positive feedback loop. To interfere with reinforced Shh signaling, we examined the potential of defined heparin and heparan sulfate (HS) polysaccharides to block Shh solubilization and Ptc receptor binding. We confirm in vitro and in vivo that proteolytic cleavage of the N-terminal Cardin-Weintraub (CW) amino acid motif is a prerequisite for Shh solubilization and function. Consistent with the established binding of soluble heparin or HS to the Shh CW target motif, both polysaccharides impaired proteolytic Shh processing and release from source cells. We also show that HS and heparin bind to, and block, another set of basic amino acids required for unimpaired Shh binding to Ptc receptors on receiving cells. Both modes of Shh activity downregulation depend more on HS size and overall charge than on specific HS sulfation modifications. We conclude that heparin oligosaccharide interference in the physiological roles of HS in Shh release and reception may be used to expand the field of investigation to pharmaceutical intervention of tumor-promoting Shh functions.
Keywords: hedgehog; heparan sulfate; heparin.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
An emerging role of Sonic hedgehog shedding as a modulator of heparan sulfate interactions.J Biol Chem. 2012 Dec 21;287(52):43708-19. doi: 10.1074/jbc.M112.356667. Epub 2012 Nov 1. J Biol Chem. 2012. PMID: 23118222 Free PMC article.
-
Kinetic and structural studies on interactions between heparin or heparan sulfate and proteins of the hedgehog signaling pathway.Biochemistry. 2007 Apr 3;46(13):3933-41. doi: 10.1021/bi6025424. Epub 2007 Mar 10. Biochemistry. 2007. PMID: 17348690 Free PMC article.
-
Sonic hedgehog processing and release are regulated by glypican heparan sulfate proteoglycans.J Cell Sci. 2015 Jun 15;128(12):2374-85. doi: 10.1242/jcs.170670. Epub 2015 May 12. J Cell Sci. 2015. PMID: 25967551
-
Interactions of signaling proteins, growth factors and other proteins with heparan sulfate: mechanisms and mysteries.Connect Tissue Res. 2015;56(4):272-80. doi: 10.3109/03008207.2015.1045066. Connect Tissue Res. 2015. PMID: 26076122 Free PMC article. Review.
-
Pseudo-active sites of protease domains: HGF/Met and Sonic hedgehog signaling in cancer.Biol Chem. 2010 Aug;391(8):881-92. doi: 10.1515/BC.2010.098. Biol Chem. 2010. PMID: 20536384 Review.
Cited by
-
Development and validation of a hedgehog heparin-binding assay for high-throughput screening.MethodsX. 2020 Dec 31;8:101207. doi: 10.1016/j.mex.2020.101207. eCollection 2021. MethodsX. 2020. PMID: 34434730 Free PMC article.
-
Two-way Dispatched function in Sonic hedgehog shedding and transfer to high-density lipoproteins.Elife. 2024 Sep 19;12:RP86920. doi: 10.7554/eLife.86920. Elife. 2024. PMID: 39297609 Free PMC article.
-
Heparanase and the hallmarks of cancer.J Transl Med. 2020 Nov 30;18(1):453. doi: 10.1186/s12967-020-02624-1. J Transl Med. 2020. PMID: 33256730 Free PMC article. Review.
-
Identification of small molecule antagonists of sonic hedgehog/heparin binding with activity in hedgehog functional assays.Biochim Biophys Acta Gen Subj. 2024 Nov;1868(11):130692. doi: 10.1016/j.bbagen.2024.130692. Epub 2024 Aug 14. Biochim Biophys Acta Gen Subj. 2024. PMID: 39151833
-
Hedgehog is relayed through dynamic heparan sulfate interactions to shape its gradient.Nat Commun. 2023 Feb 10;14(1):758. doi: 10.1038/s41467-023-36450-y. Nat Commun. 2023. PMID: 36765094 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
