

The crystal structure of dGTPase reveals the molecular basis of dGTP selectivity
- PMID: 31019074
- PMCID: PMC6511015
- DOI: 10.1073/pnas.1814999116
The crystal structure of dGTPase reveals the molecular basis of dGTP selectivity
Abstract
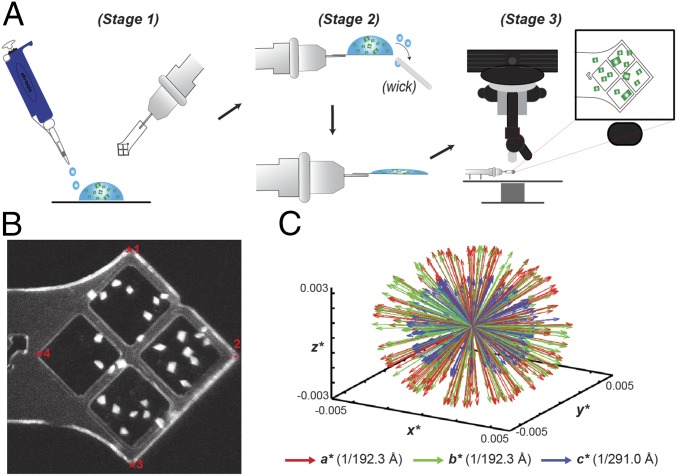
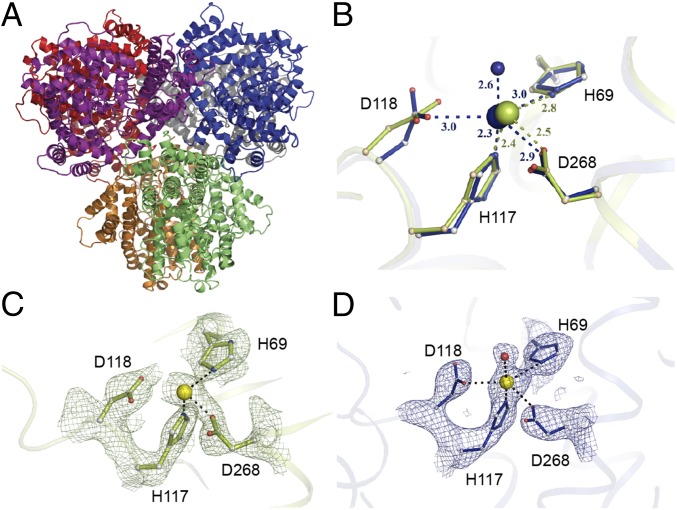
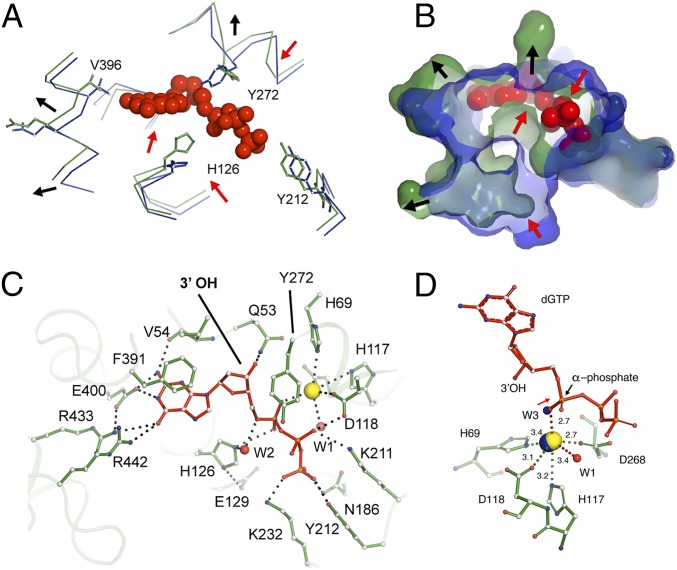
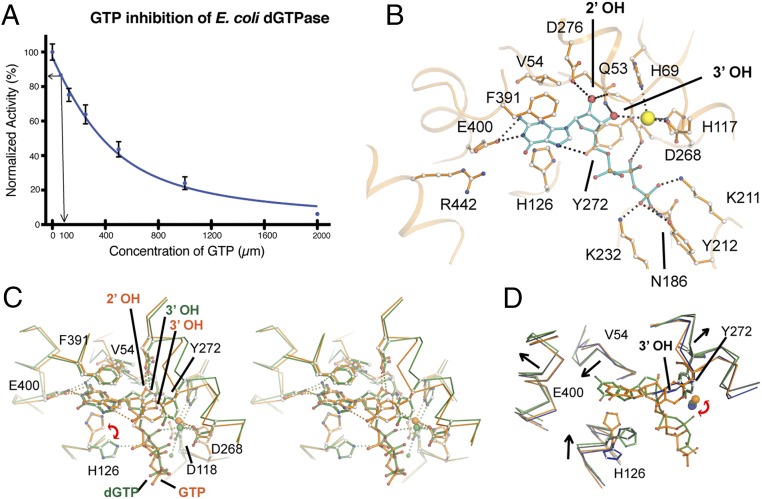
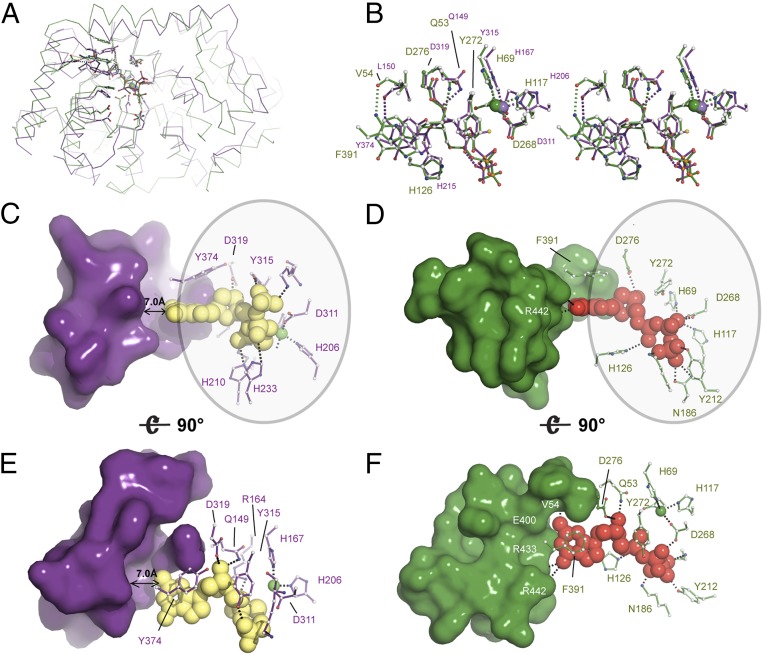
Deoxynucleotide triphosphohydrolases (dNTPases) play a critical role in cellular survival and DNA replication through the proper maintenance of cellular dNTP pools. While the vast majority of these enzymes display broad activity toward canonical dNTPs, such as the dNTPase SAMHD1 that blocks reverse transcription of retroviruses in macrophages by maintaining dNTP pools at low levels, Escherichia coli (Ec)-dGTPase is the only known enzyme that specifically hydrolyzes dGTP. However, the mechanism behind dGTP selectivity is unclear. Here we present the free-, ligand (dGTP)- and inhibitor (GTP)-bound structures of hexameric Ec-dGTPase, including an X-ray free-electron laser structure of the free Ec-dGTPase enzyme to 3.2 Å. To obtain this structure, we developed a method that applied UV-fluorescence microscopy, video analysis, and highly automated goniometer-based instrumentation to map and rapidly position individual crystals randomly located on fixed target holders, resulting in the highest indexing rates observed for a serial femtosecond crystallography experiment. Our structures show a highly dynamic active site where conformational changes are coupled to substrate (dGTP), but not inhibitor binding, since GTP locks dGTPase in its apo- form. Moreover, despite no sequence homology, Ec-dGTPase and SAMHD1 share similar active-site and HD motif architectures; however, Ec-dGTPase residues at the end of the substrate-binding pocket mimic Watson-Crick interactions providing guanine base specificity, while a 7-Å cleft separates SAMHD1 residues from dNTP bases, abolishing nucleotide-type discrimination. Furthermore, the structures shed light on the mechanism by which long distance binding (25 Å) of single-stranded DNA in an allosteric site primes the active site by conformationally "opening" a tyrosine gate allowing enhanced substrate binding.
Keywords: X-ray free-electron laser; dNTP triphosphohydrolase; metalloenzymes; serial femtosecond crystallography.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Meuth M. The genetic consequences of nucleotide precursor pool imbalance in mammalian cells. Mutat Res. 1984;126:107–112. - PubMed
-
- Seto D, Bhatnagar SK, Bessman MJ. The purification and properties of deoxyguanosine triphosphate triphosphohydrolase from Escherichia coli. J Biol Chem. 1988;263:1494–1499. - PubMed
-
- Nordlund P, Reichard P. Ribonucleotide reductases. Annu Rev Biochem. 2006;75:681–706. - PubMed
-
- Kunz BA, et al. International commission for protection against environmental mutagens and carcinogens. Deoxyribonucleoside triphosphate levels: A critical factor in the maintenance of genetic stability. Mutat Res. 1994;318:1–64. - PubMed
-
- Wheeler LJ, Rajagopal I, Mathews CK. Stimulation of mutagenesis by proportional deoxyribonucleoside triphosphate accumulation in Escherichia coli. DNA Repair (Amst) 2005;4:1450–1456. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
