

Overlooked roles of DNA damage and maternal age in generating human germline mutations
- PMID: 31019089
- PMCID: PMC6511033
- DOI: 10.1073/pnas.1901259116
Overlooked roles of DNA damage and maternal age in generating human germline mutations
Abstract
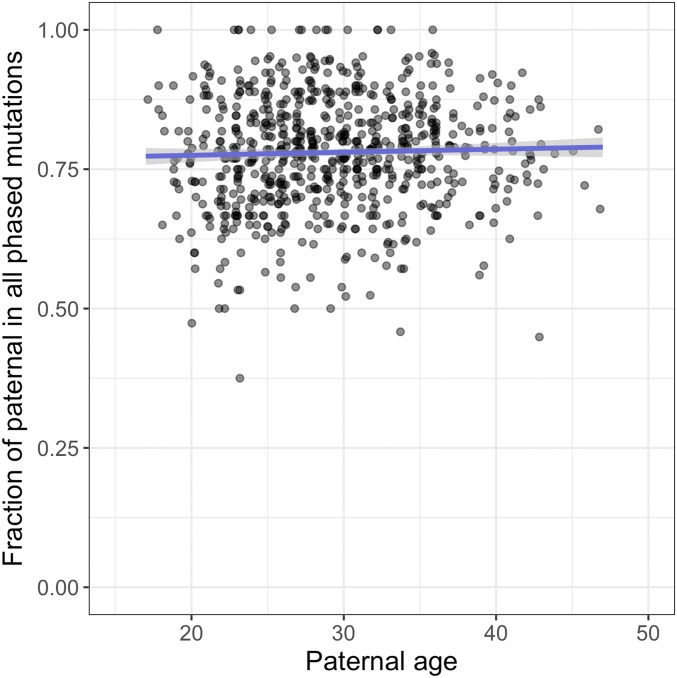
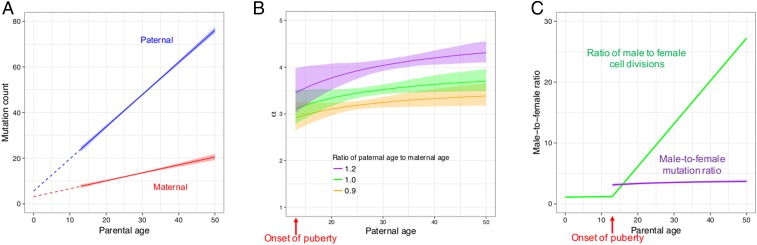
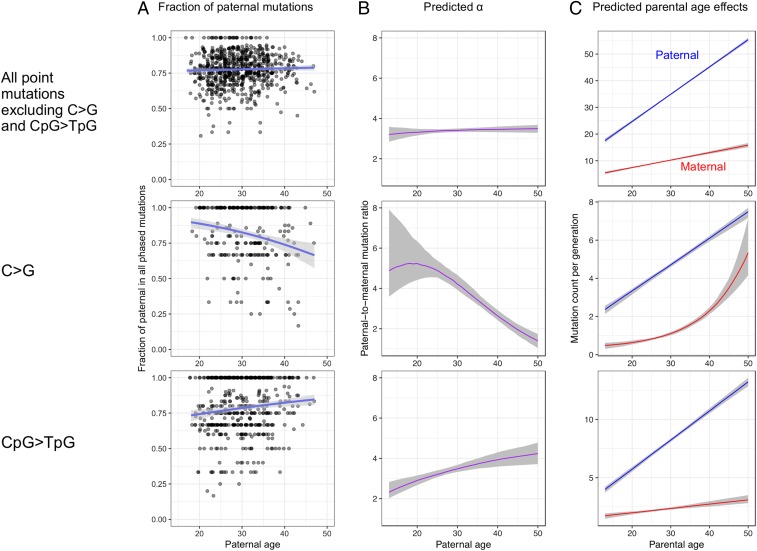
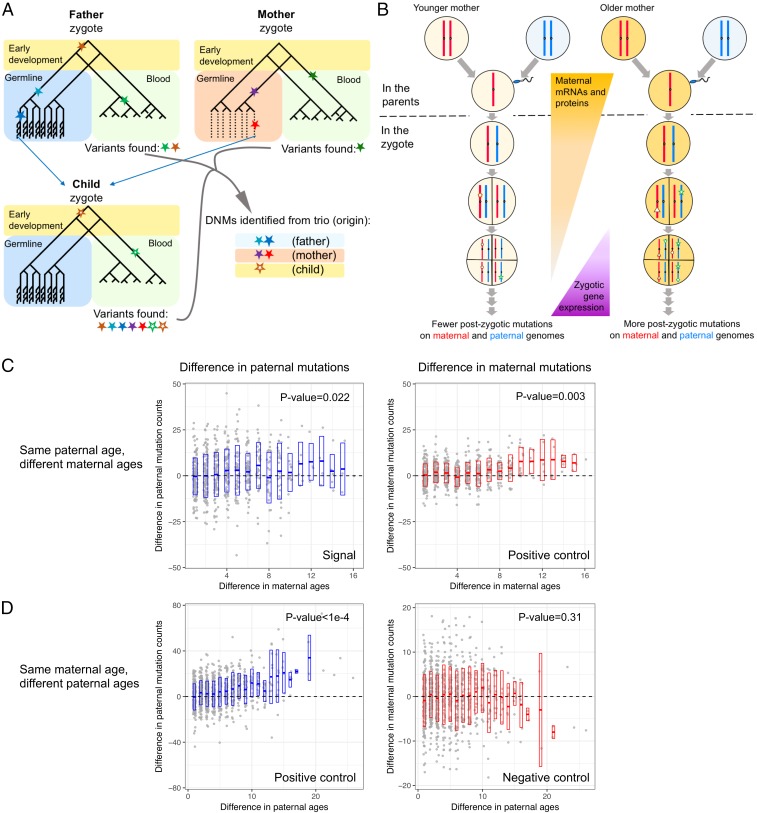
The textbook view that most germline mutations in mammals arise from replication errors is indirectly supported by the fact that there are both more mutations and more cell divisions in the male than in the female germline. When analyzing large de novo mutation datasets in humans, we find multiple lines of evidence that call that view into question. Notably, despite the drastic increase in the ratio of male to female germ cell divisions after the onset of spermatogenesis, even young fathers contribute three times more mutations than young mothers, and this ratio barely increases with parental age. This surprising finding points to a substantial contribution of damage-induced mutations. Indeed, C-to-G transversions and CpG transitions, which together constitute over one-fourth of all base substitution mutations, show genomic distributions and sex-specific age dependencies indicative of double-strand break repair and methylation-associated damage, respectively. Moreover, we find evidence that maternal age at conception influences the mutation rate both because of the accumulation of damage in oocytes and potentially through an influence on the number of postzygotic mutations in the embryo. These findings reveal underappreciated roles of DNA damage and maternal age in the genesis of human germline mutations.
Keywords: DNA damage and repair; DNA replication; germline mutation; male mutation bias; maternal age effect.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Huttley GA, Jakobsen IB, Wilson SR, Easteal S. How important is DNA replication for mutagenesis? Mol Biol Evol. 2000;17:929–937. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
