

Peptone-mediated glucagon-like peptide-1 secretion depends on intestinal absorption and activation of basolaterally located Calcium-Sensing Receptors
- PMID: 31020803
- PMCID: PMC6482282
- DOI: 10.14814/phy2.14056
Peptone-mediated glucagon-like peptide-1 secretion depends on intestinal absorption and activation of basolaterally located Calcium-Sensing Receptors
Abstract
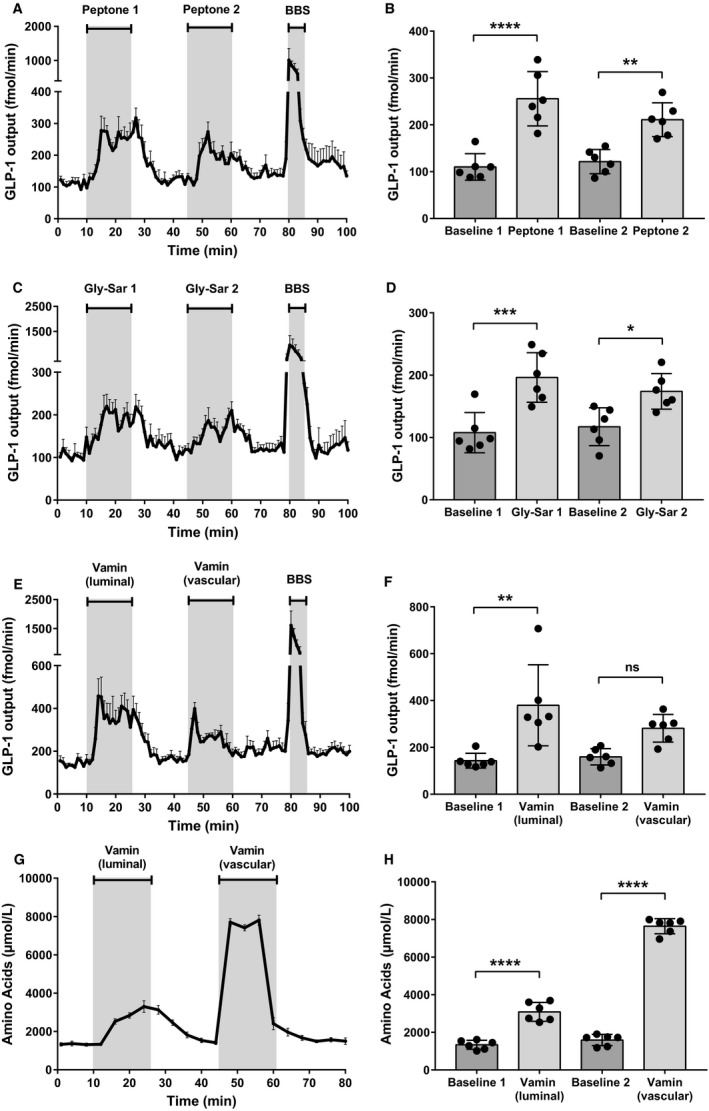
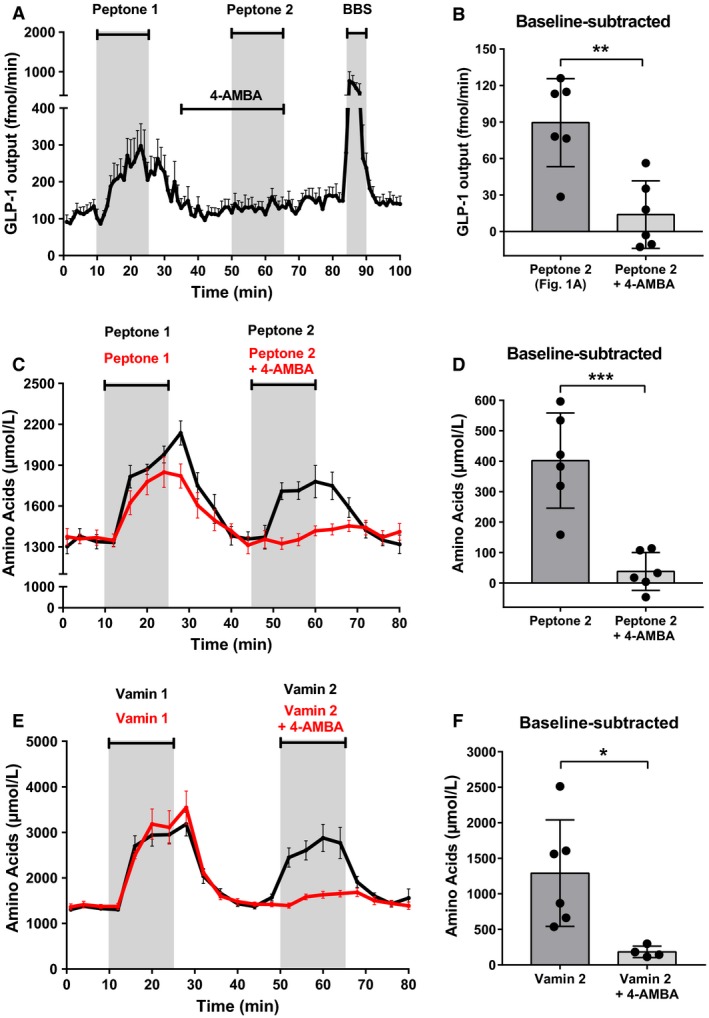
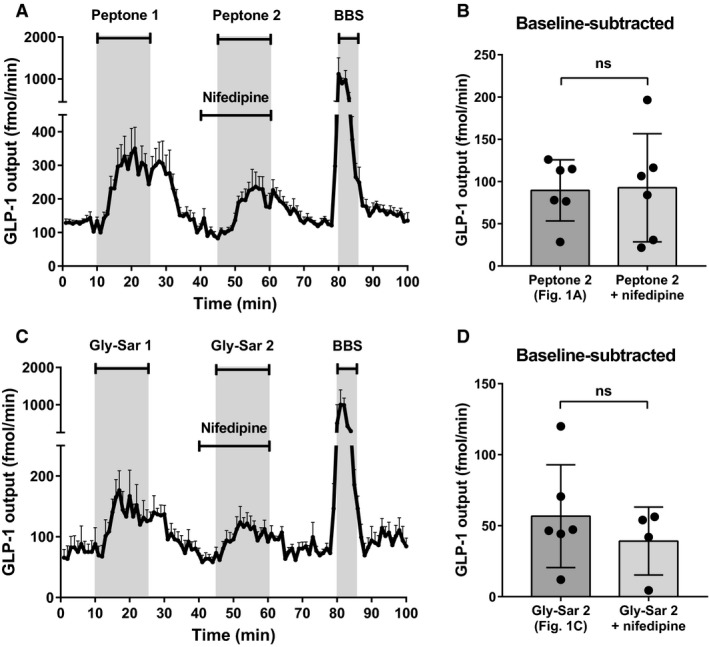
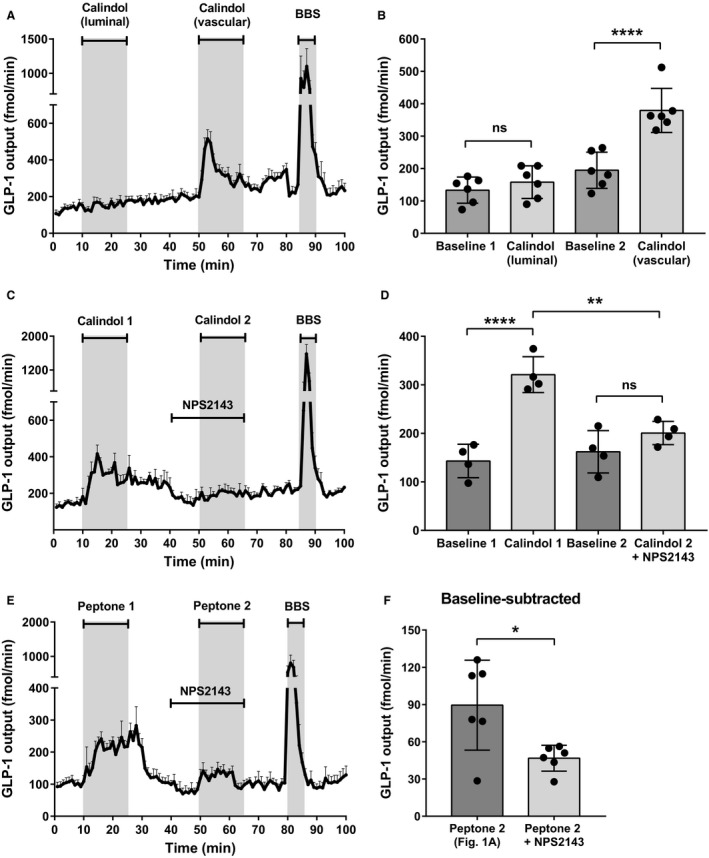
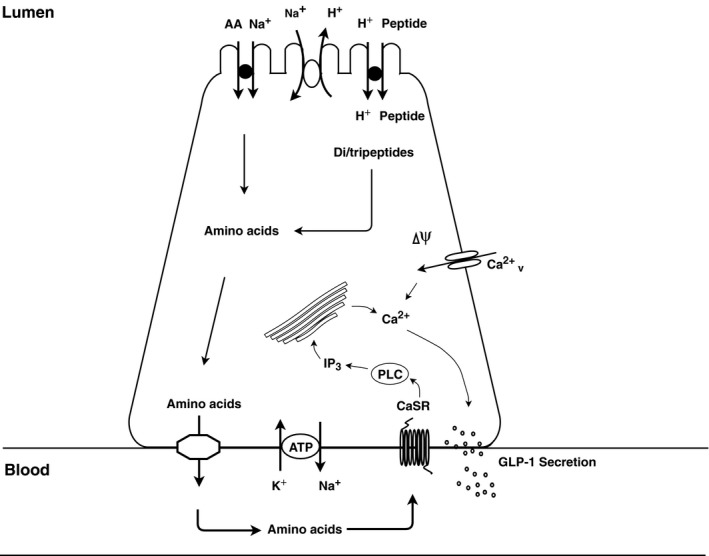
Protein intake robustly stimulates the secretion of the incretin hormone, glucagon-like peptide-1 (GLP-1) but the molecular mechanisms involved are not well understood. In particular, it is unknown whether proteins stimulate secretion by activation of luminal or basolateral sensors. We characterized the mechanisms using a physiologically relevant model - the isolated perfused proximal rat small intestine. Intraluminal protein hydrolysates derived from meat (peptone; 50 mg/mL) increased GLP-1 secretion 2.3-fold (from a basal secretion of 110 ± 28 fmol/min). The sensory mechanisms underlying the response depended on di/tripeptide uptake through Peptide Transporter 1 (PepT1) and subsequent basolateral activation of the amino acid sensing receptor, Calcium-Sensing Receptor (CaSR), since inhibition of PepT1 as well as CaSR both attenuated the peptone-induced GLP-1 response. Supporting this, intraluminal peptones were absorbed efficiently by the perfused intestine (resulting in increased amino acid concentrations in the venous effluent) and infusion of amino acids robustly stimulated GLP-1 secretion. Inhibitors of voltage-gated L-type Ca2+ channels had no effect on secretion suggesting that peptone-mediated GLP-1 secretion is not mediated by L-cell depolarization with subsequent opening of these channels. Specific targeting of CaSR could serve as a target to stimulate the endogenous secretion of GLP-1.
Keywords: Amino acid sensing; Calcium-Sensing Receptor; glucagon-like peptide 1; peptide transporter 1; peptone.
© 2019 The Authors. Physiological Reports published by Wiley Periodicals, Inc. on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
The authors of this work declare no potential conflicts of interest relevant to this article.
Figures
References
-
- Baggio, L. L. , and Drucker D. J.. 2007. Biology of incretins: GLP‐1 and GIP. Gastroenterology 132:2131–2157. - PubMed
-
- Belza, A. , Ritz C., Sørensen M. Q., Holst J. J., Rehfeld J. F., and Astrup A.. 2013. Contribution of gastroenteropancreatic appetite hormones to protein‐induced satiety. Am. J. Clin. Nutr. 97:980–989. - PubMed
-
- Clemmensen, C. , Jørgensen C. V., Smajilovic S., and Bräuner‐Osborne H.. 2017. Robust GLP‐1 secretion by basic L‐amino acids does not require the GPRC6A receptor. Diabetes Obes. Metab. 19:599–603. - PubMed
-
- Conigrave, A. D. , and Hampson D. R.. 2006. Broad‐spectrum l‐amino acid sensing by class 3 G‐protein‐coupled receptors. Trends Endocrinol. Metab. 17:398–407. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
