

Convergent evolution of hetero-oligomeric cellulose synthesis complexes in mosses and seed plants
- PMID: 31021018
- PMCID: PMC6711812
- DOI: 10.1111/tpj.14366
Convergent evolution of hetero-oligomeric cellulose synthesis complexes in mosses and seed plants
Abstract
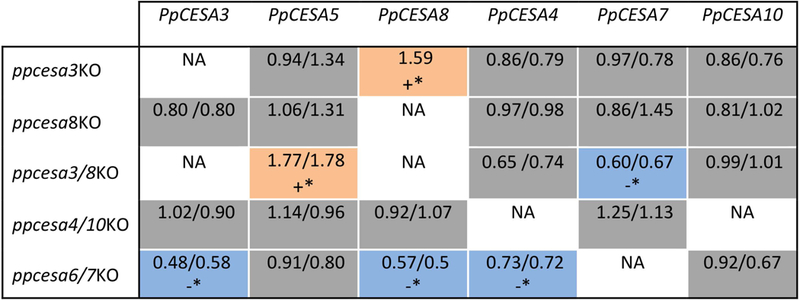
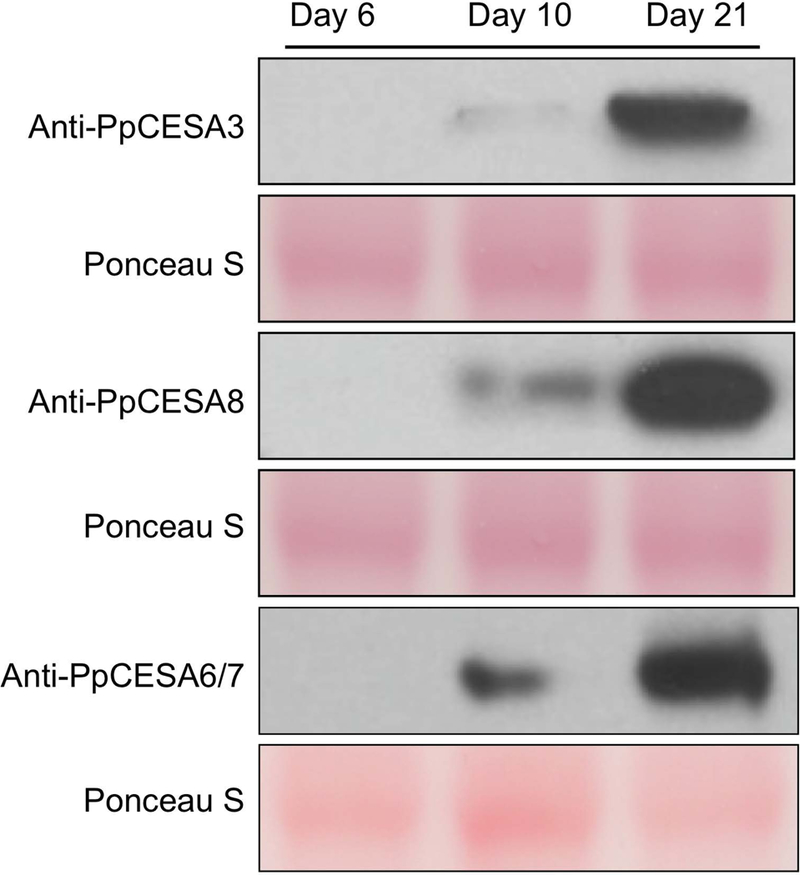
In seed plants, cellulose is synthesized by rosette-shaped cellulose synthesis complexes (CSCs) that are obligate hetero-oligomeric, comprising three non-interchangeable cellulose synthase (CESA) isoforms. The moss Physcomitrella patens has rosette CSCs and seven CESAs, but its common ancestor with seed plants had rosette CSCs and a single CESA gene. Therefore, if P. patens CSCs are hetero-oligomeric, then CSCs of this type evolved convergently in mosses and seed plants. Previous gene knockout and promoter swap experiments showed that PpCESAs from class A (PpCESA3 and PpCESA8) and class B (PpCESA6 and PpCESA7) have non-redundant functions in secondary cell wall cellulose deposition in leaf midribs, whereas the two members of each class are redundant. Based on these observations, we proposed the hypothesis that the secondary class A and class B PpCESAs associate to form hetero-oligomeric CSCs. Here we show that transcription of secondary class A PpCESAs is reduced when secondary class B PpCESAs are knocked out and vice versa, as expected for genes encoding isoforms that occupy distinct positions within the same CSC. The class A and class B isoforms co-accumulate in developing gametophores and co-immunoprecipitate, suggesting that they interact to form a complex in planta. Finally, secondary PpCESAs interact with each other, whereas three of four fail to self-interact when expressed in two different heterologous systems. These results are consistent with the hypothesis that obligate hetero-oligomeric CSCs evolved independently in mosses and seed plants and we propose the constructive neutral evolution hypothesis as a plausible explanation for convergent evolution of hetero-oligomeric CSCs.
Keywords: Physcomitrella patens; cell wall; cellulose; cellulose synthase; cellulose synthesis complex; convergent evolution.
© 2019 The Authors The Plant Journal © 2019 John Wiley & Sons Ltd.
Conflict of interest statement
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Figures






References
-
- Arioli T, Peng L, Betzner AS, Burn J, Wittke W, Herth W, Camilleri C, Höfte H, Plazinski J, Birch R, Cork A, Glover J, Redmond J and Williamson RE (1998) Molecular analysis of cellulose biosynthesis in Arabidopsis. Science, 279, 717–720. - PubMed
-
- Atanassov II, Pittman JK and Turner SR (2009) Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J Biol Chem, 284, 3833–3841. - PubMed
-
- Betancur L, Singh B, Rapp RA, Wendel JF, Marks MD, Roberts AW and Haigler CH (2011) Phylogenetically distinct cellulose synthase genes support secondary wall thickening in arabidopsis shoot trichomes and cotton fiber. J Integr Plant Biol, 52, 205–220. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
