

Classification and Nomenclature of CRISPR-Cas Systems: Where from Here?
- PMID: 31021272
- PMCID: PMC6636873
- DOI: 10.1089/crispr.2018.0033
Classification and Nomenclature of CRISPR-Cas Systems: Where from Here?
Abstract
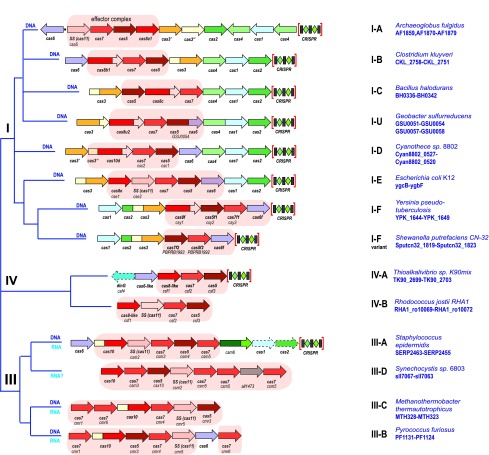
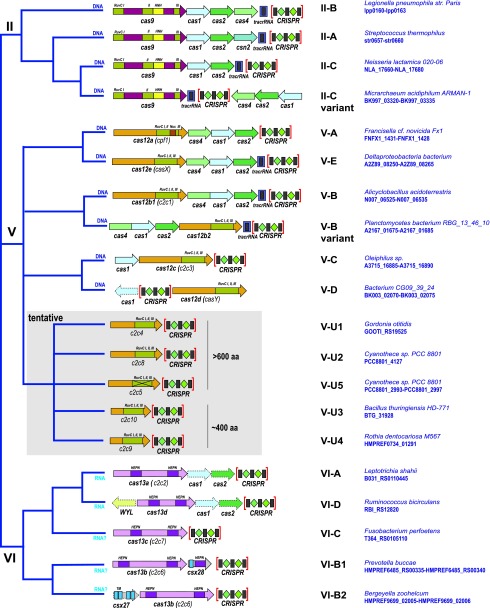
As befits an immune mechanism, CRISPR-Cas systems are highly variable with respect to Cas protein sequences, gene composition, and organization of the genomic loci. Optimal classification of CRISPR-Cas systems and rational nomenclature for CRISPR-associated genes are essential for further progress of CRISPR research. These are highly challenging tasks because of the complexity of CRISPR-Cas and their fast evolution, including frequent module shuffling, as well as the lack of universal markers for a consistent evolutionary classification. The complexity and variability of CRISPR-Cas systems necessitate a multipronged approach to classification and nomenclature. We present a brief summary of the current state of the art and discuss further directions in this area.
Conflict of interest statement
No competing financial interests exist.
Figures



References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
