

TXNIP mediates high glucose-induced mitophagic flux and lysosome enlargement in human retinal pigment epithelial cells
- PMID: 31023645
- PMCID: PMC6503994
- DOI: 10.1242/bio.038521
TXNIP mediates high glucose-induced mitophagic flux and lysosome enlargement in human retinal pigment epithelial cells
Erratum in
-
Correction: TXNIP mediates high glucose-induced mitophagic flux and lysosome enlargement in human retinal pigment epithelial cells.Biol Open. 2022 Aug 15;11(8):bio059489. doi: 10.1242/bio.059489. Epub 2022 Jul 29. Biol Open. 2022. PMID: 35904988 Free PMC article. No abstract available.
Abstract
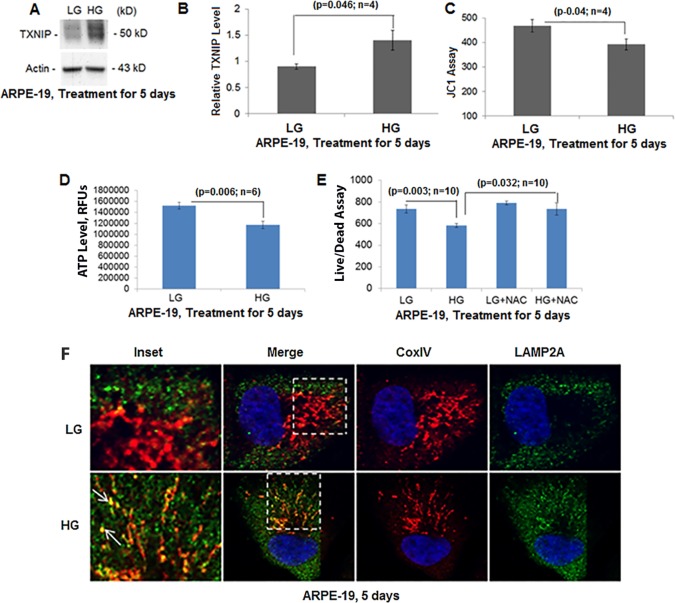
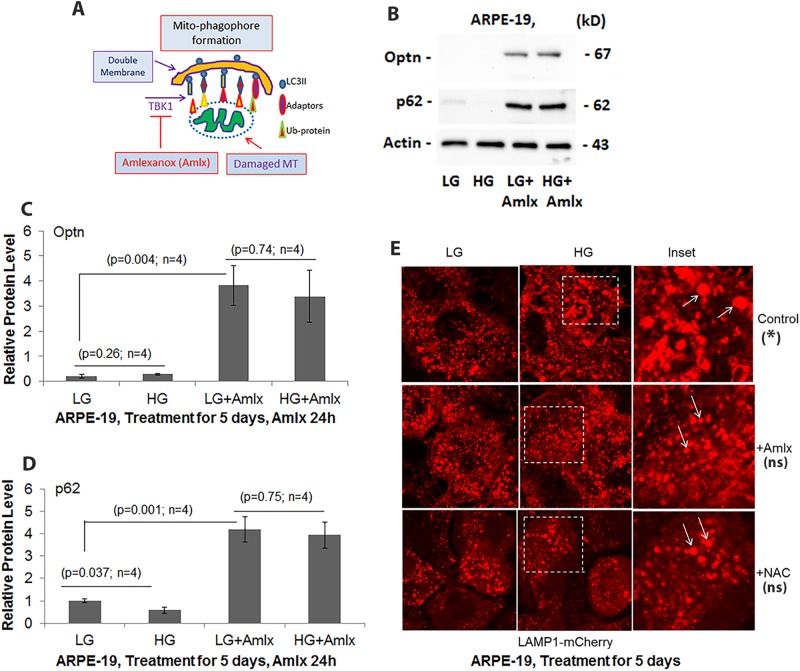
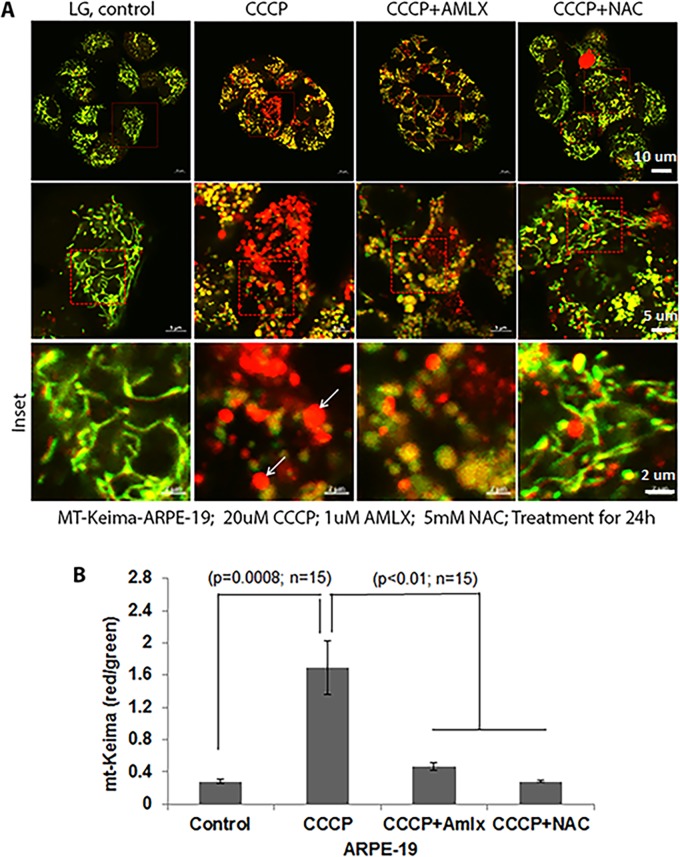
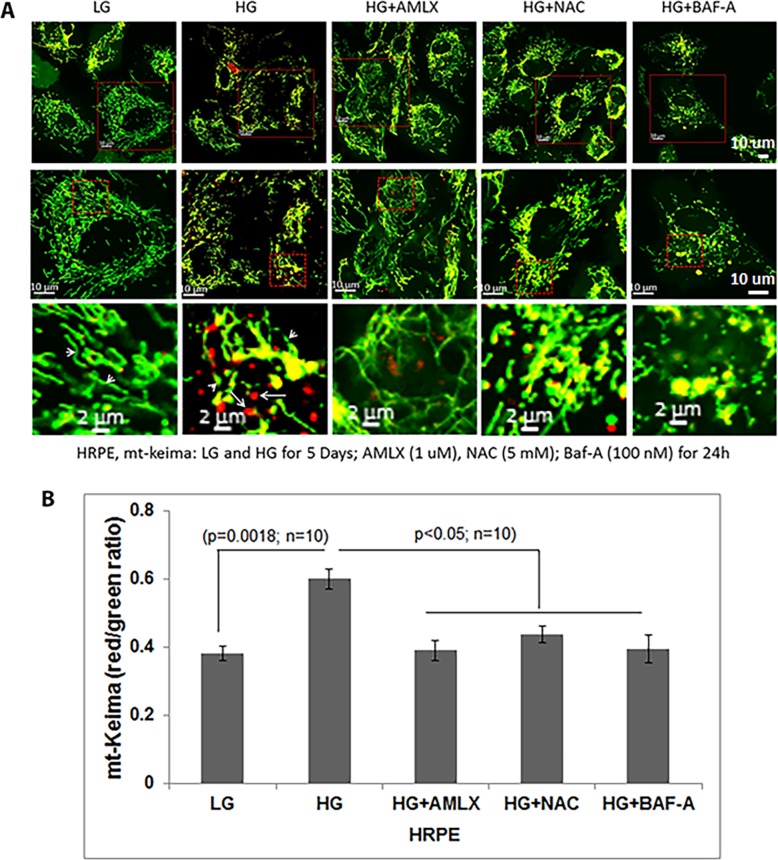
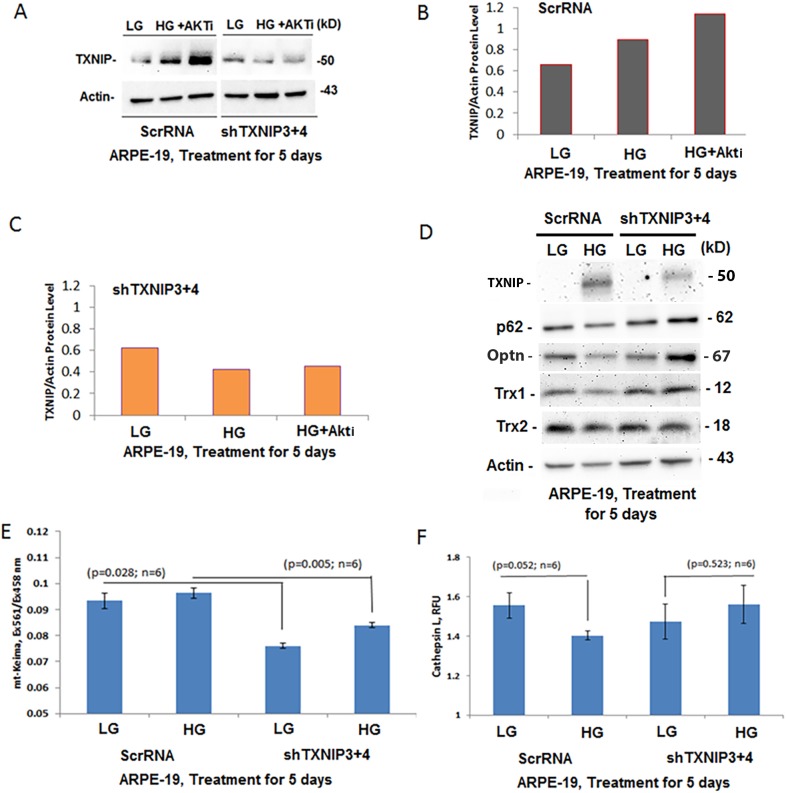
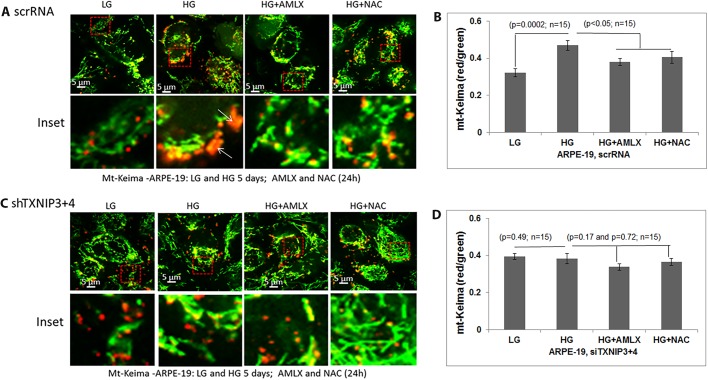
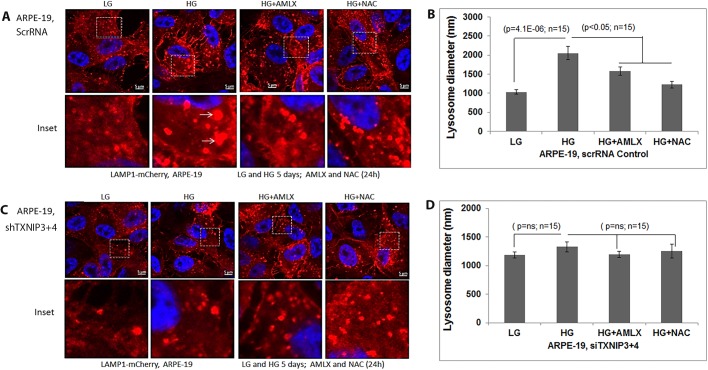
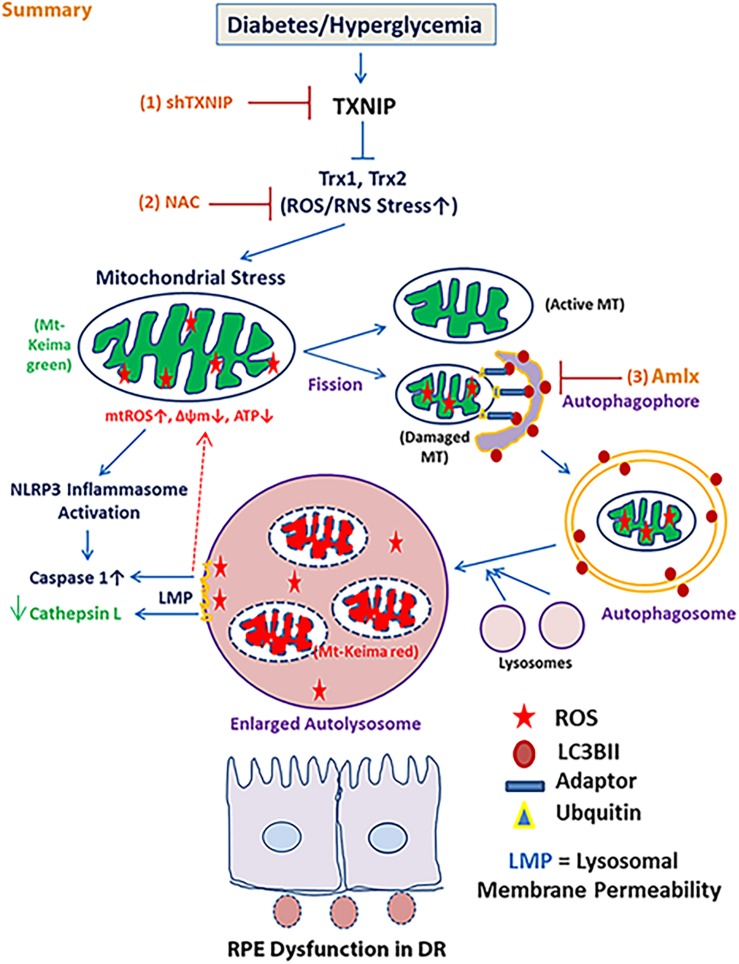
Thioredoxin-interacting protein (TXNIP) plays a critical role in oxidative stress, inflammation, apoptosis and the pathogenesis of diabetic retinopathy (DR). However, the role of TXNIP in high glucose-induced retinal pigment epithelium (RPE) dysfunction is still unknown. Here, we show that high glucose (HG; 25 mM,) significantly increases TXNIP expression at both the mRNA and protein levels when compared to low glucose (LG; 5.5 mM) in a human RPE cell line (ARPE-19) and primary human RPE (HRPE) cells. TXNIP upregulation is associated with mitochondrial membrane depolarization, fragmentation and mitophagic flux to lysosomes. We used confocal live-cell imaging of RPE cells expressing mt-Keima, a coral protein that emits green light in mitochondria (alkaline or neutral pH) and red light in the acidic lysosome, to measure mitophagic flux. We observed an elongated mitochondrial network of green mt-Keima under LG, which is fragmented in HG. Red mt-Keima accumulates in lysosomes as small punctate aggregations under LG in both ARPE-19 and HRPE cells, whereas they are significantly enlarged (two- to threefold) under HG. Lysosomal enlargement under HG is further illustrated by lysosomal membrane protein LAMP1-mCherry expression in both ARPE-19 and HRPE cells. Furthermore, HG causes lysosomal cathepsin L inactivation and pro-inflammatory caspase-1 activation in ARPE-19 cells. TXNIP knockdown by shRNA prevents mitochondrial fragmentation, mitophagic flux and lysosome enlargement under HG. In addition, antioxidant N-acetylcysteine (NAC) and Amlexanox (Amlx), an inhibitor of protein kinase TBK1 and of the mitophagic adaptors Optineurin (Optn) and Sequestosome 1 (p62/SQSTM1), prevent mitophagic flux and lysosome enlargement. These results suggest that TXNIP mediates several deleterious effects of high glucose on RPE, which may be implicated in the development of DR.
Keywords: Hyperglycemia; Lysosome destabilization; Mitophagy; Retinal pigment epithelium; TXNIP.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Bodur C., Kazyken D., Huang K., Ekim Ustunel B., Siroky K. A., Tooley A. S., Gonzalez I. E., Foley D. H., Acosta-Jaquez H. A., Barnes T. M. et al. (2018). The IKK-related kinase TBK1 activates mTORC1 directly in response to growth factors and innate immune agonists. EMBO J. 37, 19-38. 10.15252/embj.201696164 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
