

METTL1 Promotes let-7 MicroRNA Processing via m7G Methylation
- PMID: 31031083
- PMCID: PMC6591002
- DOI: 10.1016/j.molcel.2019.03.040
METTL1 Promotes let-7 MicroRNA Processing via m7G Methylation
Abstract
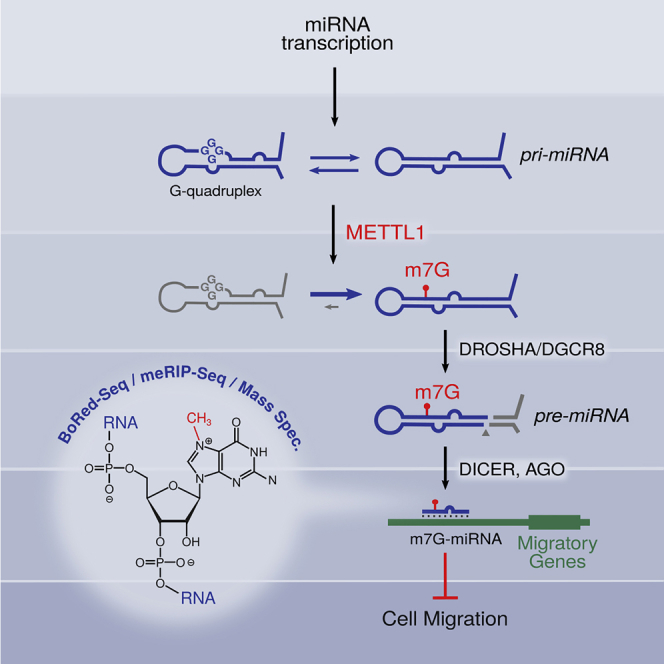
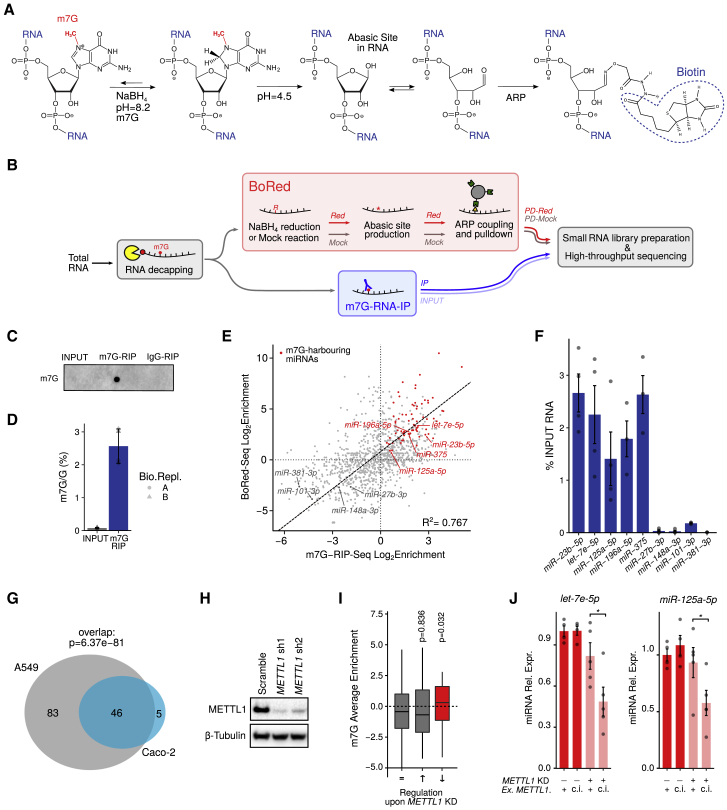
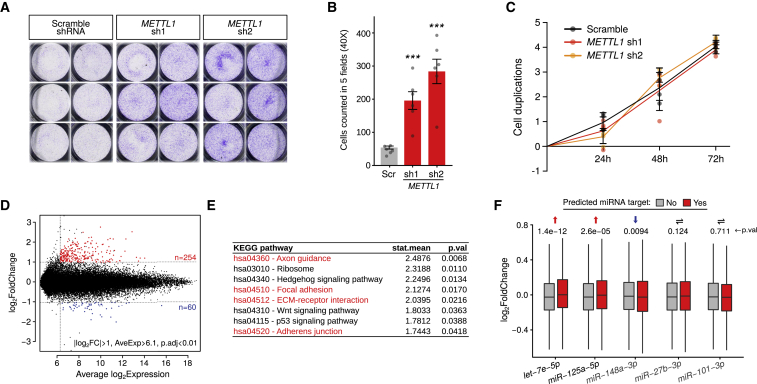
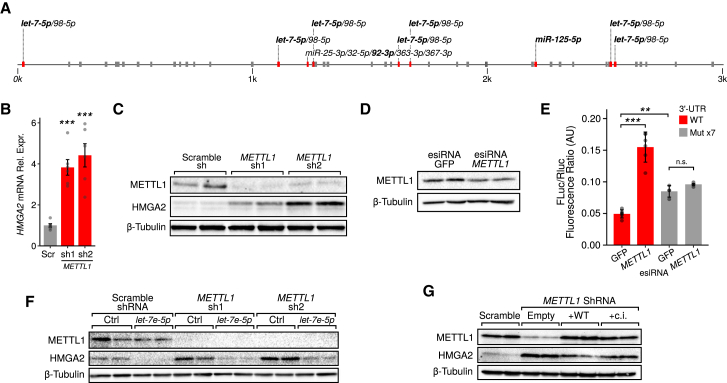
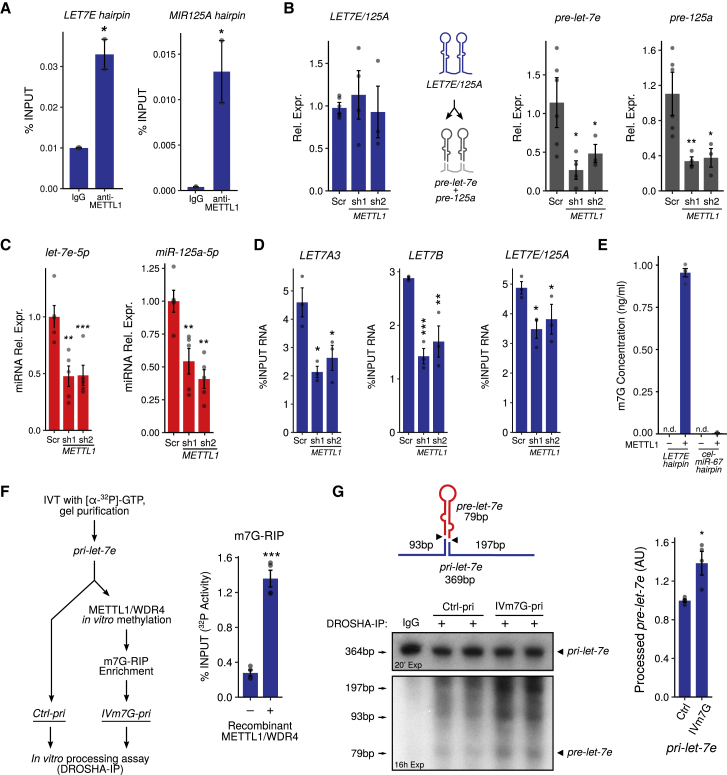
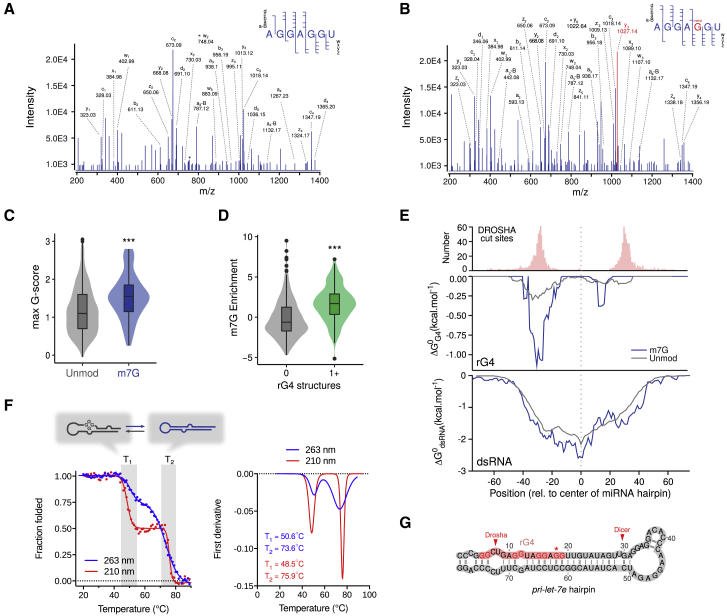
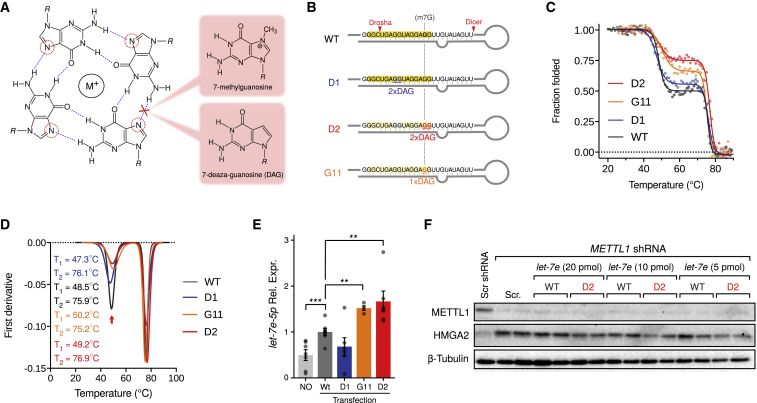
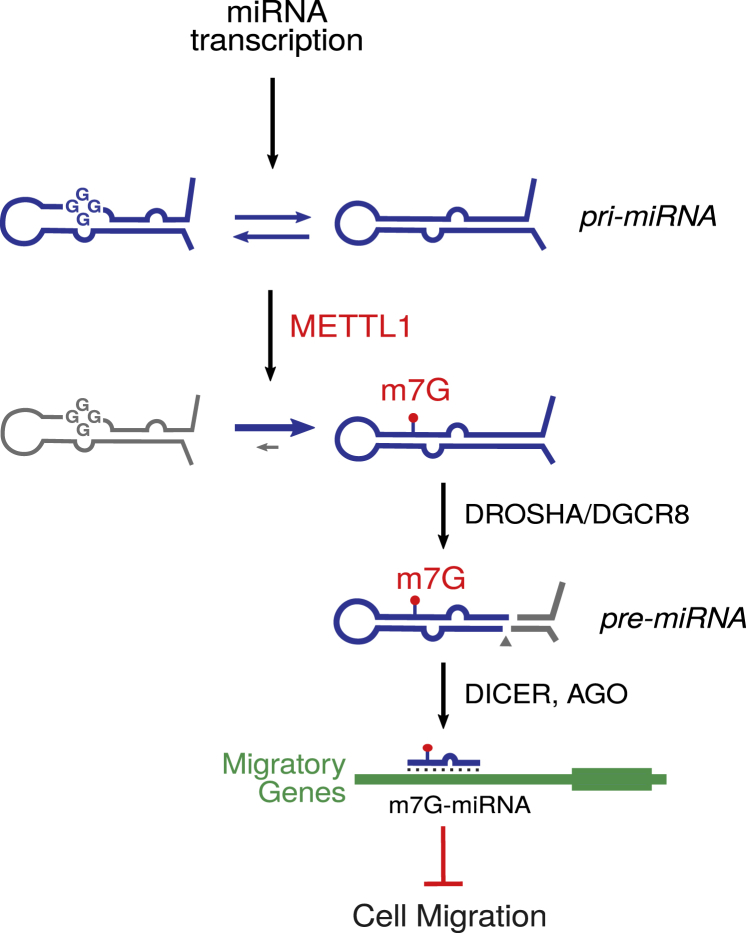
7-methylguanosine (m7G) is present at mRNA caps and at defined internal positions within tRNAs and rRNAs. However, its detection within low-abundance mRNAs and microRNAs (miRNAs) has been hampered by a lack of sensitive detection strategies. Here, we adapt a chemical reactivity assay to detect internal m7G in miRNAs. Using this technique (Borohydride Reduction sequencing [BoRed-seq]) alongside RNA immunoprecipitation, we identify m7G within a subset of miRNAs that inhibit cell migration. We show that the METTL1 methyltransferase mediates m7G methylation within miRNAs and that this enzyme regulates cell migration via its catalytic activity. Using refined mass spectrometry methods, we map m7G to a single guanosine within the let-7e-5p miRNA. We show that METTL1-mediated methylation augments let-7 miRNA processing by disrupting an inhibitory secondary structure within the primary miRNA transcript (pri-miRNA). These results identify METTL1-dependent N7-methylation of guanosine as a new RNA modification pathway that regulates miRNA structure, biogenesis, and cell migration.
Keywords: 7-methylguanosine; G-quadruplexes; METTL1; RNA methylation; SAM-dependent methyltransferase; cell migration; high-throughput sequencing; let-7; miRNA biogenesis; microRNA.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Put the Pedal to the METTL1: Adding Internal m7G Increases mRNA Translation Efficiency and Augments miRNA Processing.Mol Cell. 2019 Jun 20;74(6):1105-1107. doi: 10.1016/j.molcel.2019.06.004. Mol Cell. 2019. PMID: 31226274
-
No Evidence for N7-Methylation of Guanosine (m7G) in Human let-7e.Mol Cell. 2020 Jul 16;79(2):199-200. doi: 10.1016/j.molcel.2020.05.022. Mol Cell. 2020. PMID: 32679072 No abstract available.
-
Further Evidence Supporting N7-Methylation of Guanosine (m7G) in Human MicroRNAs.Mol Cell. 2020 Jul 16;79(2):201-202. doi: 10.1016/j.molcel.2020.05.023. Mol Cell. 2020. PMID: 32679073 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
