

Cholesterol Induces CD8+ T Cell Exhaustion in the Tumor Microenvironment
- PMID: 31031094
- PMCID: PMC7061417
- DOI: 10.1016/j.cmet.2019.04.002
Cholesterol Induces CD8+ T Cell Exhaustion in the Tumor Microenvironment
Abstract
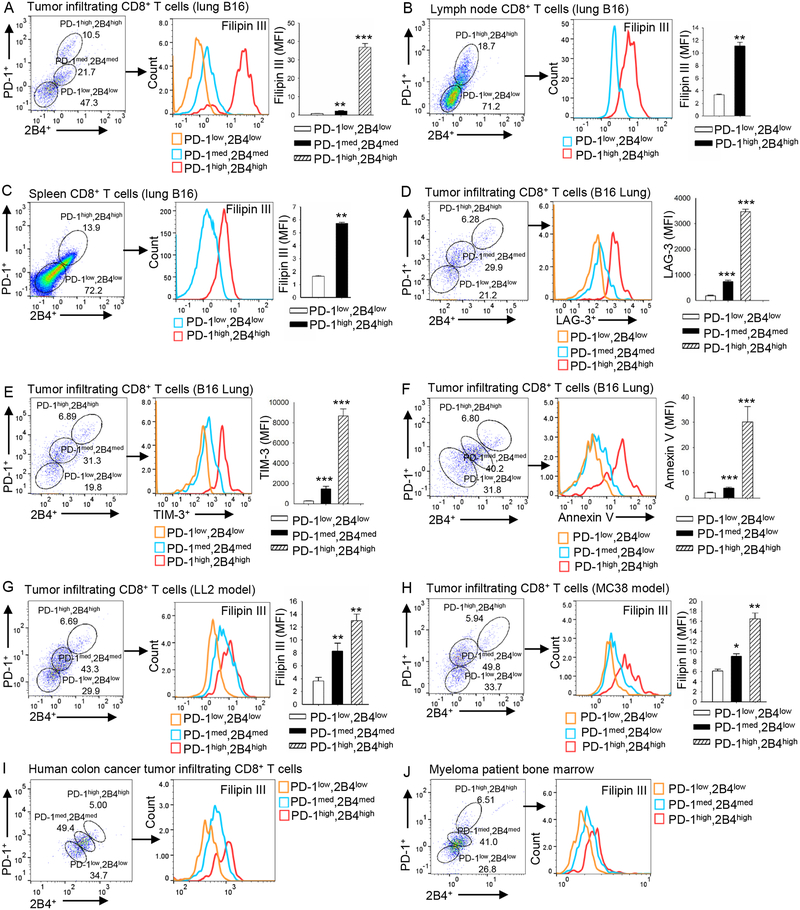
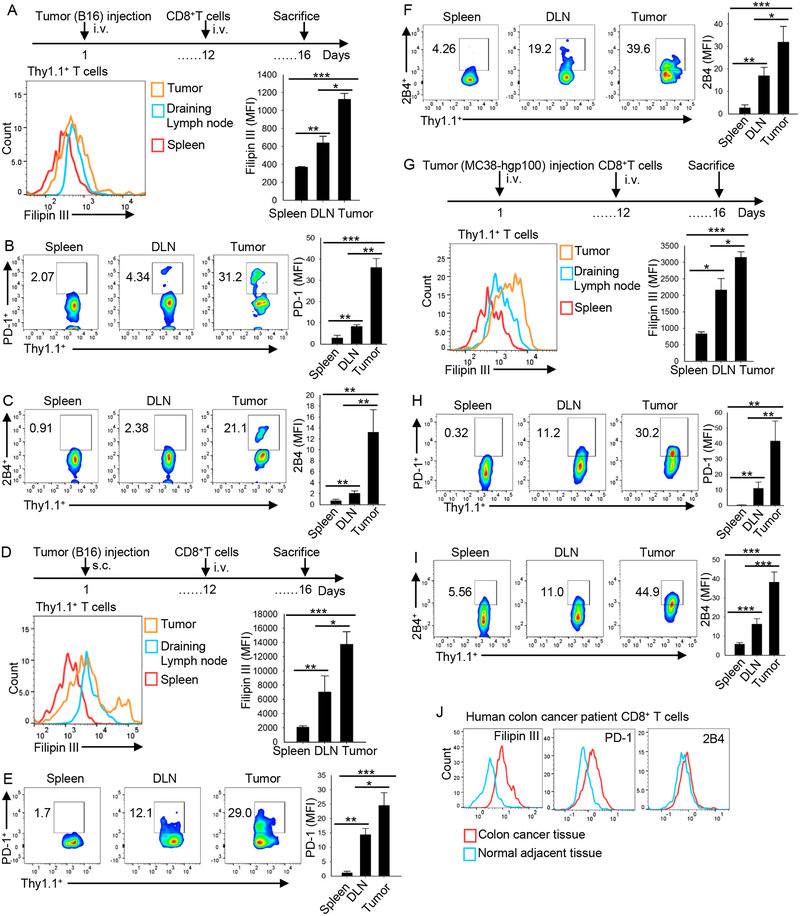
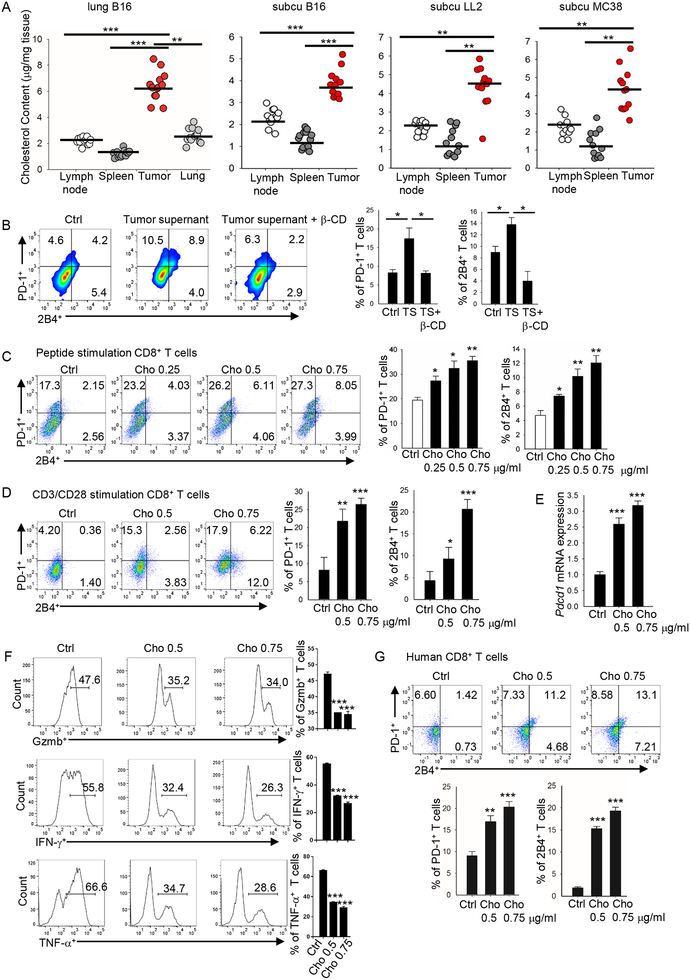
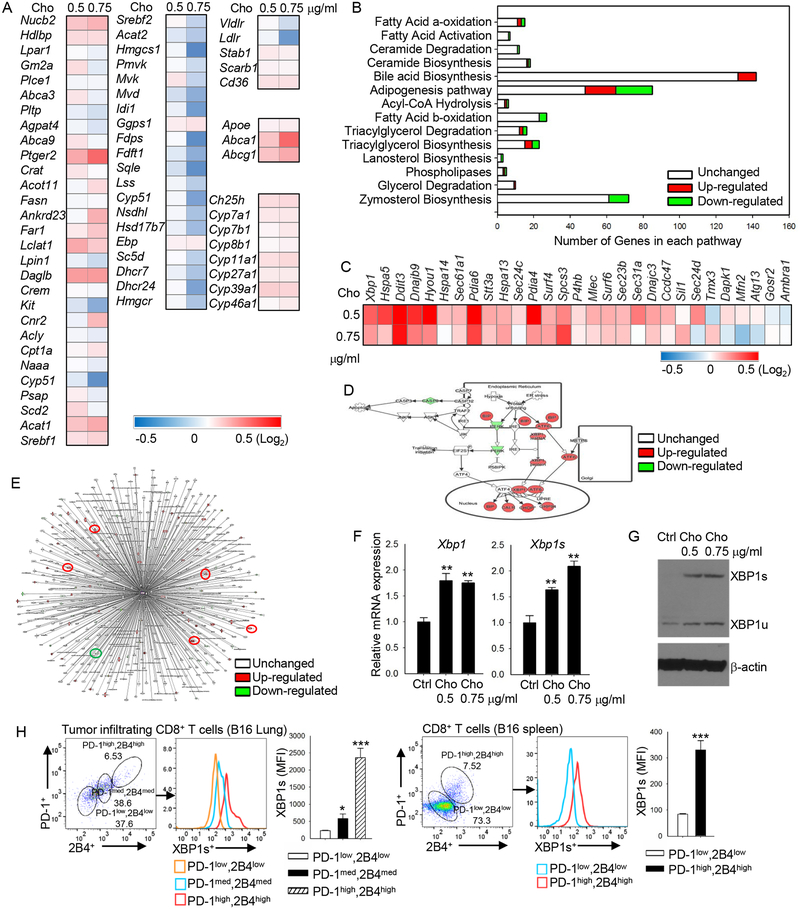
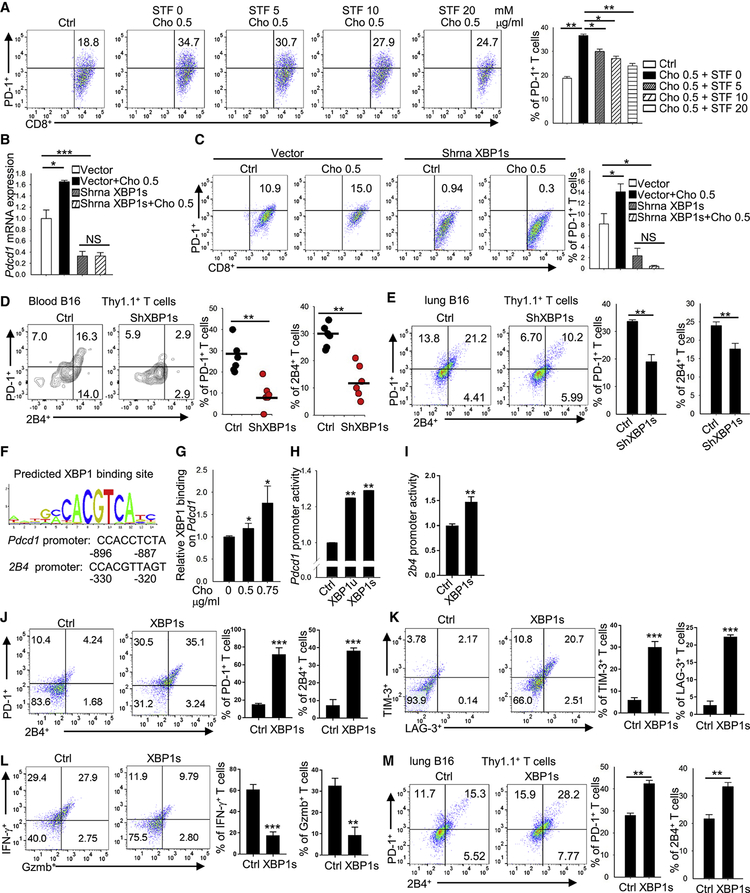
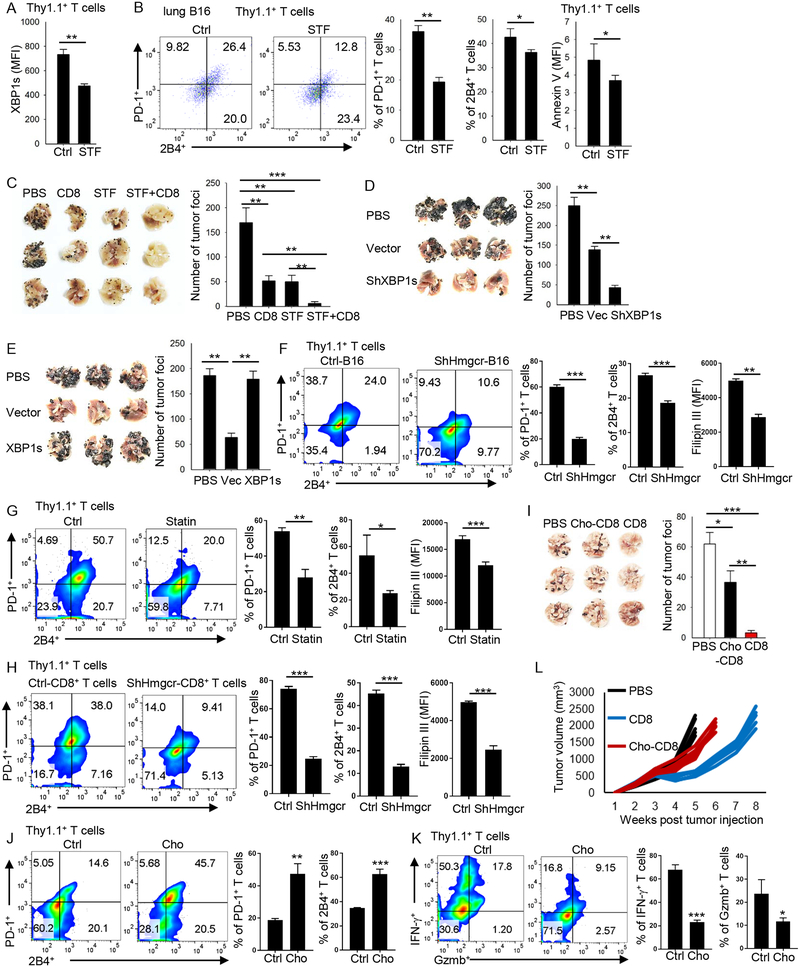
Tumor-infiltrating T cells often lose their effector function; however, the mechanisms are incompletely understood. We report that cholesterol in the tumor microenvironment induces CD8+ T cell expression of immune checkpoints and exhaustion. Tumor tissues enriched with cholesterol and cholesterol content in tumor-infiltrating CD8+ T cells were positively and progressively associated with upregulated T cell expression of PD-1, 2B4, TIM-3, and LAG-3. Adoptively transferred CD8+ T cells acquired cholesterol, expressed high levels of immune checkpoints, and became exhausted upon entering a tumor. Tumor culture supernatant or cholesterol induced immune checkpoint expression by increasing endoplasmic reticulum (ER) stress in CD8+ T cells. Consequently, the ER stress sensor XBP1 was activated and regulated PD-1 and 2B4 transcription. Inhibiting XBP1 or reducing cholesterol in CD8+ T cells effectively restored antitumor activity. This study reveals a mechanism underlying T cell exhaustion and suggests a new strategy for restoring T cell function by reducing cholesterol to enhance T cell-based immunotherapy.
Keywords: CD8+ T cells; cholesterol; exhaustion; immune checkpoints; tumor microenvironment.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors have declared that no conflicts of interest exist.
Figures
Comment in
-
Tumor Cholesterol Up, T Cells Down.Cell Metab. 2019 Jul 2;30(1):12-13. doi: 10.1016/j.cmet.2019.06.007. Cell Metab. 2019. PMID: 31269422
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
