

A Stable Visual World in Primate Primary Visual Cortex
- PMID: 31031112
- PMCID: PMC6519108
- DOI: 10.1016/j.cub.2019.03.069
A Stable Visual World in Primate Primary Visual Cortex
Abstract
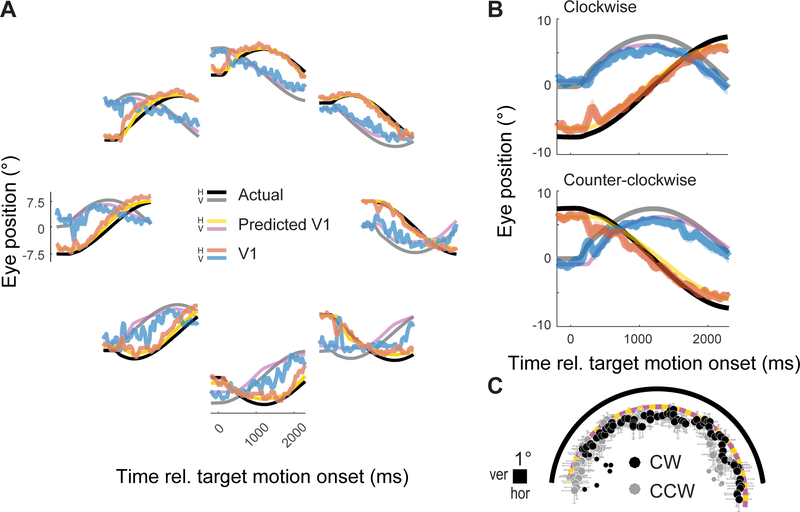
Humans and other primates rely on eye movements to explore visual scenes and to track moving objects. As a result, the image that is projected onto the retina-and propagated throughout the visual cortical hierarchy-is almost constantly changing and makes little sense without taking into account the momentary direction of gaze. How is this achieved in the visual system? Here, we show that in primary visual cortex (V1), the earliest stage of cortical vision, neural representations carry an embedded "eye tracker" that signals the direction of gaze associated with each image. Using chronically implanted multi-electrode arrays, we recorded the activity of neurons in area V1 of macaque monkeys during tasks requiring fast (exploratory) and slow (pursuit) eye movements. Neurons were stimulated with flickering, full-field luminance noise at all times. As in previous studies, we observed neurons that were sensitive to gaze direction during fixation, despite comparable stimulation of their receptive fields. We trained a decoder to translate neural activity into metric estimates of gaze direction. This decoded signal tracked the eye accurately not only during fixation but also during fast and slow eye movements. After a fast eye movement, the eye-position signal arrived in V1 at approximately the same time at which the new visual information arrived from the retina. Using simulations, we show that this V1 eye-position signal could be used to take into account the sensory consequences of eye movements and map the fleeting positions of objects on the retina onto their stable position in the world.
Keywords: computation; electrophysiology; eye position; population coding; primary visual cortex; vision.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing financial interests.
Figures





Similar articles
-
Encoding of smooth pursuit direction and eye position by neurons of area MSTd of macaque monkey.J Neurosci. 1997 May 15;17(10):3847-60. doi: 10.1523/JNEUROSCI.17-10-03847.1997. J Neurosci. 1997. PMID: 9133403 Free PMC article.
-
Gaze-dependent visual neurons in area V3A of monkey prestriate cortex.J Neurosci. 1989 Apr;9(4):1112-25. doi: 10.1523/JNEUROSCI.09-04-01112.1989. J Neurosci. 1989. PMID: 2703870 Free PMC article.
-
Visual receptive fields of neurons in primary visual cortex (V1) move in space with the eye movements of fixation.Vision Res. 1997 Feb;37(3):257-65. doi: 10.1016/s0042-6989(96)00182-4. Vision Res. 1997. PMID: 9135859
-
The neural basis of smooth pursuit eye movements in the rhesus monkey brain.Brain Cogn. 2008 Dec;68(3):229-40. doi: 10.1016/j.bandc.2008.08.014. Epub 2008 Oct 2. Brain Cogn. 2008. PMID: 18835077 Review.
-
A physiological perspective on fixational eye movements.Vision Res. 2016 Jan;118:31-47. doi: 10.1016/j.visres.2014.12.006. Epub 2014 Dec 20. Vision Res. 2016. PMID: 25536465 Free PMC article. Review.
Cited by
-
Movement-Related Signals in Sensory Areas: Roles in Natural Behavior.Trends Neurosci. 2020 Aug;43(8):581-595. doi: 10.1016/j.tins.2020.05.005. Epub 2020 Jun 22. Trends Neurosci. 2020. PMID: 32580899 Free PMC article. Review.
-
Joint coding of visual input and eye/head position in V1 of freely moving mice.Neuron. 2022 Dec 7;110(23):3897-3906.e5. doi: 10.1016/j.neuron.2022.08.029. Epub 2022 Sep 21. Neuron. 2022. PMID: 36137549 Free PMC article.
-
Low-Level Visual Information Is Maintained across Saccades, Allowing for a Postsaccadic Handoff between Visual Areas.J Neurosci. 2020 Dec 2;40(49):9476-9486. doi: 10.1523/JNEUROSCI.1169-20.2020. Epub 2020 Oct 28. J Neurosci. 2020. PMID: 33115930 Free PMC article.
-
Activity in primate visual cortex is minimally driven by spontaneous movements.Nat Neurosci. 2023 Nov;26(11):1953-1959. doi: 10.1038/s41593-023-01459-5. Epub 2023 Oct 12. Nat Neurosci. 2023. PMID: 37828227 Free PMC article.
-
A computational account of transsaccadic attentional allocation based on visual gain fields.Proc Natl Acad Sci U S A. 2024 Jul 2;121(27):e2316608121. doi: 10.1073/pnas.2316608121. Epub 2024 Jun 28. Proc Natl Acad Sci U S A. 2024. PMID: 38941277 Free PMC article.
References
-
- Gur M, and Snodderly DM (1997). Visual receptive fields of neurons in primary visual cortex (V1) move in space with the eye movements of fixation. Vision Res. 3, 257–265. - PubMed
-
- Andersen RA, and Buneo CA (2002). Intentional maps in posterior parietal cortex. Annu. Rev. Neurosci 189–220. - PubMed
-
- Colby CL, and Goldberg ME (1999). Space and attention in parietal cortex. Annu. Rev. Neurosci 319–349. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
