

Different Hydrophobins of Fusarium graminearum Are Involved in Hyphal Growth, Attachment, Water-Air Interface Penetration and Plant Infection
- PMID: 31031728
- PMCID: PMC6474331
- DOI: 10.3389/fmicb.2019.00751
Different Hydrophobins of Fusarium graminearum Are Involved in Hyphal Growth, Attachment, Water-Air Interface Penetration and Plant Infection
Abstract
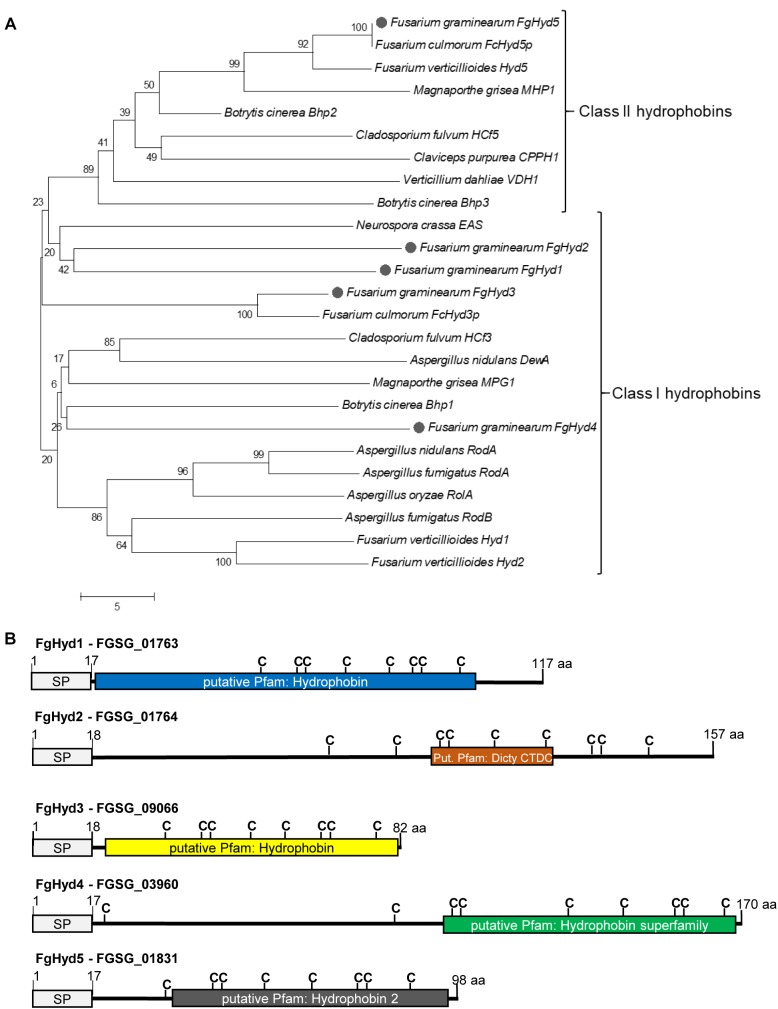
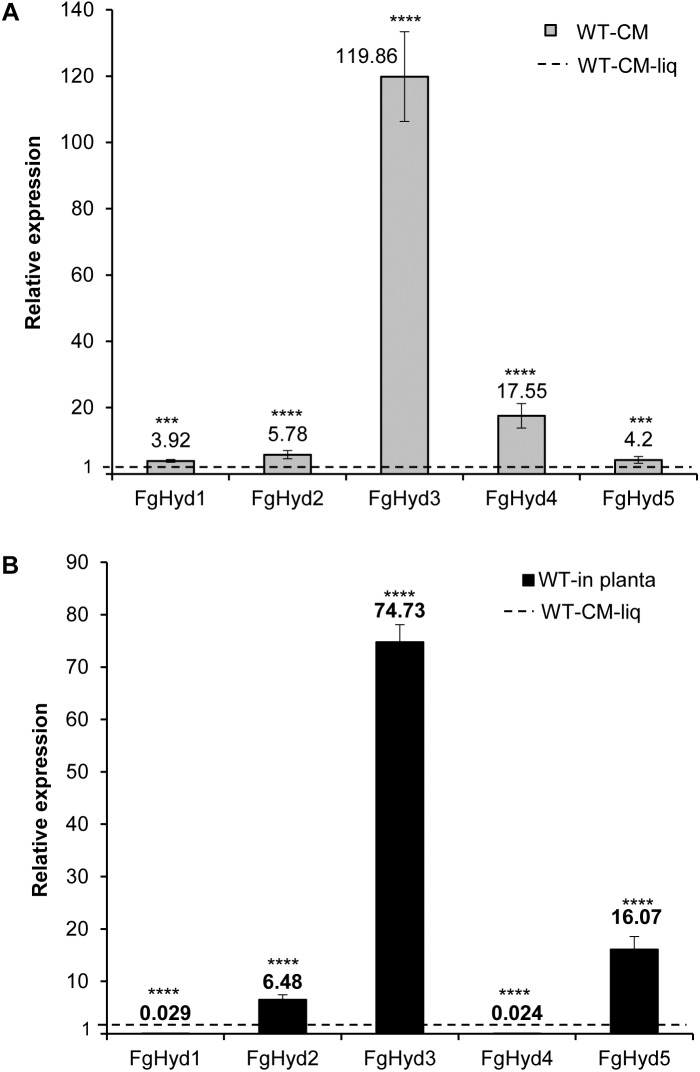
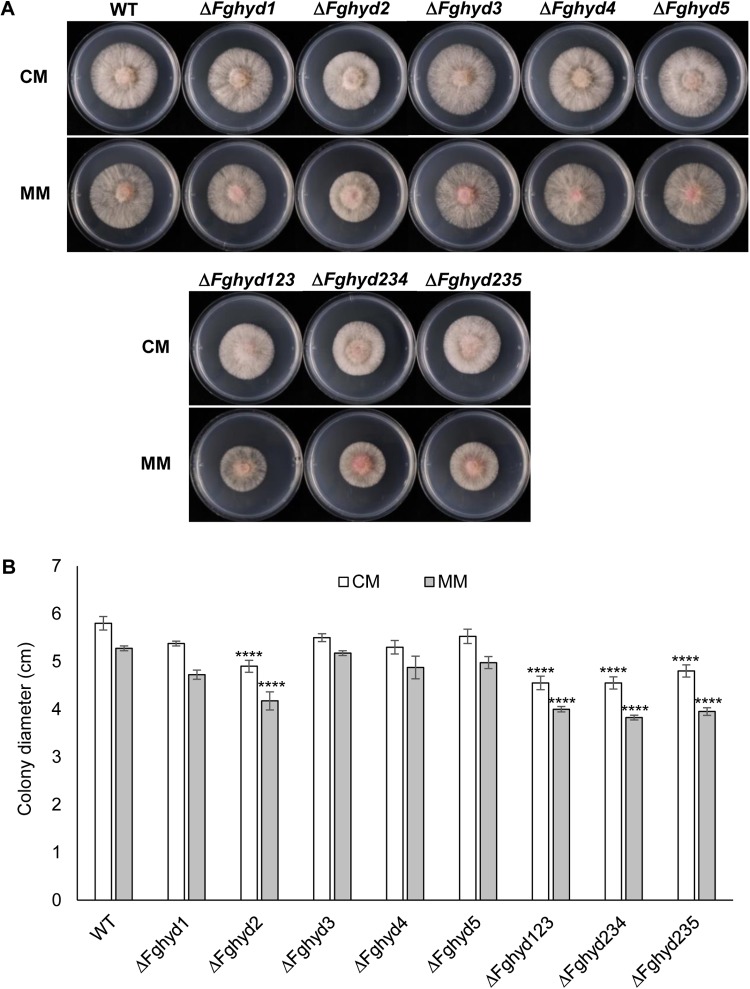
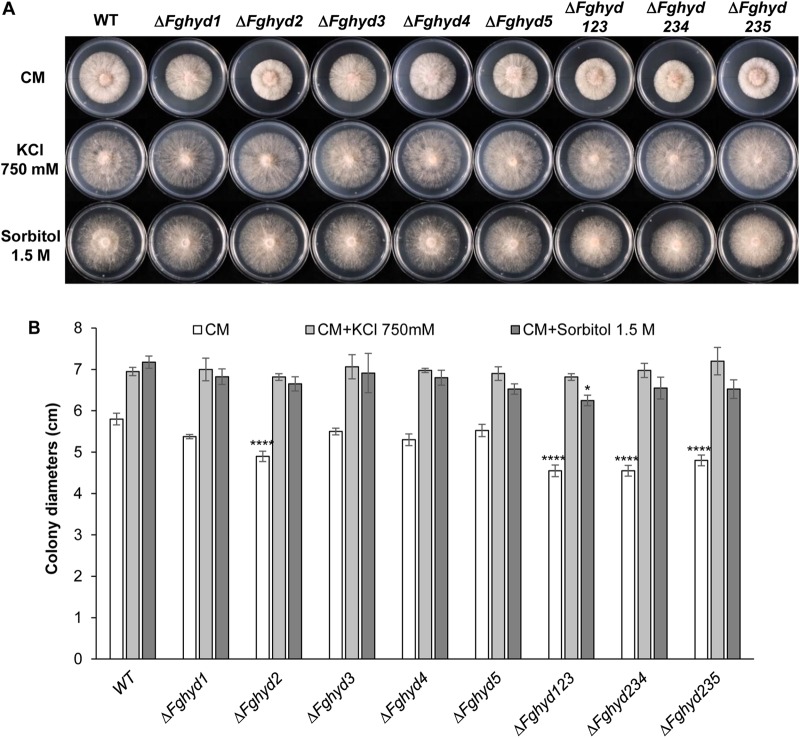
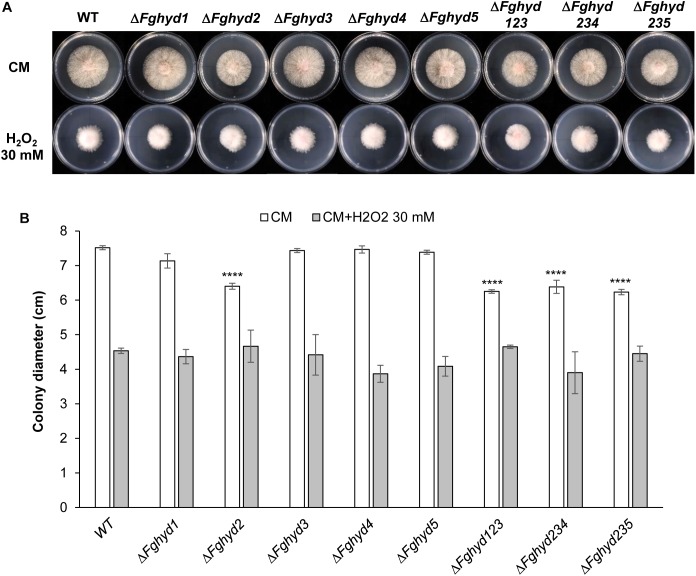
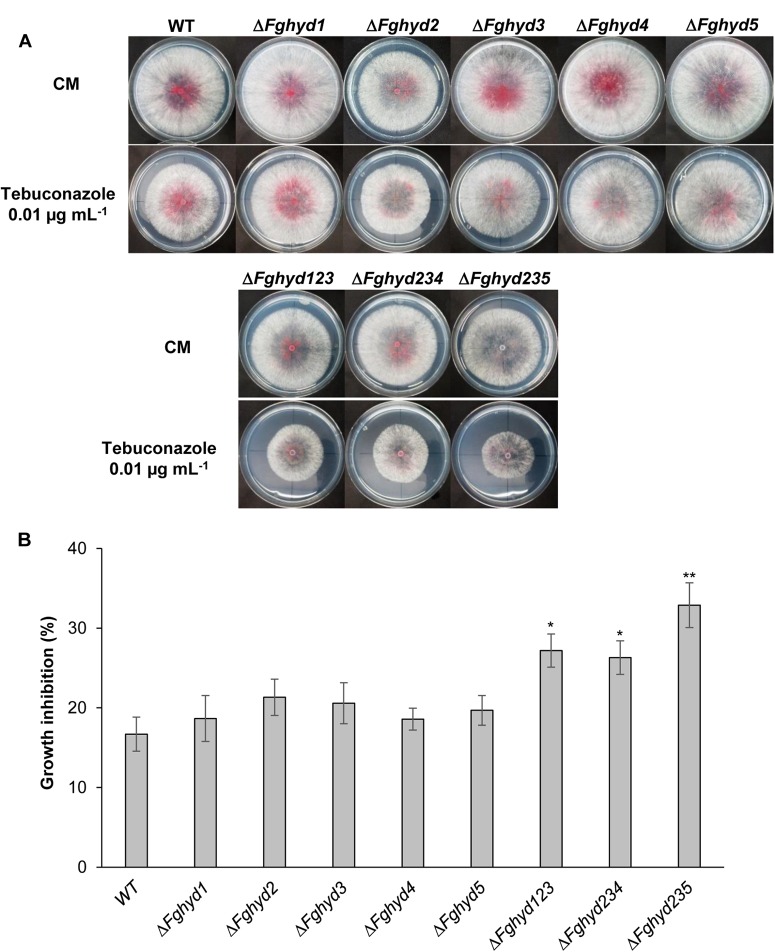
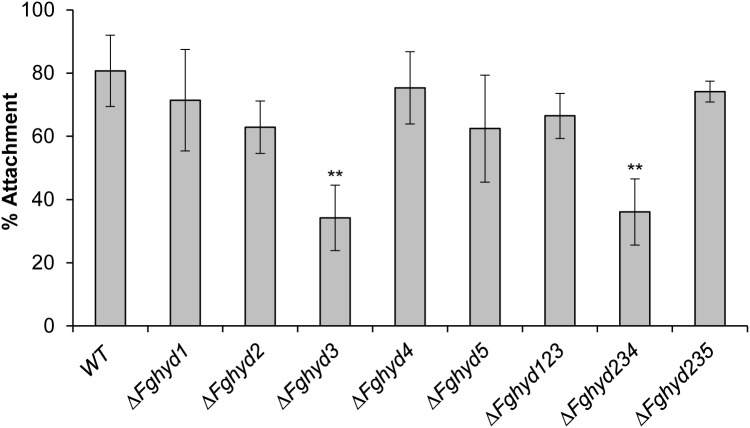
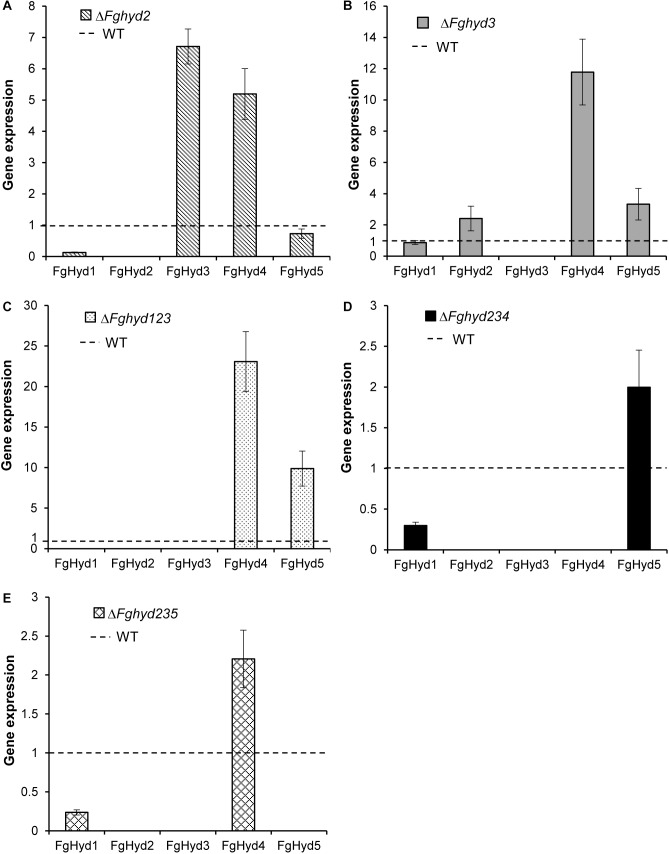
Hydrophobins (HPs) are small secreted fungal proteins possibly involved in several processes such as formation of fungal aerial structures, attachment to hydrophobic surfaces, interaction with the environment and protection against the host defense system. The genome of the necrotrophic plant pathogen Fusarium graminearum contains five genes encoding for HPs (FgHyd1-5). Single and triple FgHyd mutants were produced and characterized. A reduced growth was observed when the ΔFghyd2 and the three triple mutants including the deletion of FgHyd2 were grown in complete or minimal medium. Surprisingly, the growth of these mutants was similar to wild-type when grown under ionic, osmotic or oxidative stress conditions. All the mutant strains confirmed the ability to develop conidia and perithecia, suggesting that the FgHyds are not involved in normal development of asexual and sexual structures. A reduction in the ability of hyphae to penetrate through the water-air interface was observed for the single mutants ΔFghyd2 and ΔFghyd3 as well as for the triple mutants including the deletion of FgHyd2 and FgHyd3. Besides, ΔFghyd3 and the triple mutant ΔFghyd234 were also affected in the attachment to hydrophobic surface. Indeed, wheat infection experiments showed a reduction of symptomatic spikelets for ΔFghyd2 and ΔFghyd3 and the triple mutants only when spray inoculation was performed. This result could be ascribed to the affected ability of mutants deleted of FgHyd2 and FgHyd3 to penetrate through the water-air interface and to attach to hydrophobic surfaces such as the spike tissue. This hypothesis is strengthened by a histological analysis, performed by fluorescence microscopy, showing no defects in the morphology of infection structures produced by mutant strains. Interestingly, triple hydrophobin mutants were significantly more inhibited than wild-type by the treatment with a systemic triazole fungicide, while no defects at the cell wall level were observed.
Keywords: Fusarium graminearum; attachment; hydrophobin; virulence; water-air interface.
Figures
References
-
- Beck E., Ludwig G., Auerswald E. A., Reiss B., Schaller H. (1982). Nucleotide sequence and exact localization of the neomycin phosphotransferase gene from transposon Tn5. Gene 19 327–336. - PubMed
LinkOut - more resources
Full Text Sources