

Severe acute respiratory syndrome coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC
- PMID: 31034780
- PMCID: PMC6662968
- DOI: 10.1096/fj.201802418R
Severe acute respiratory syndrome coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC
Abstract
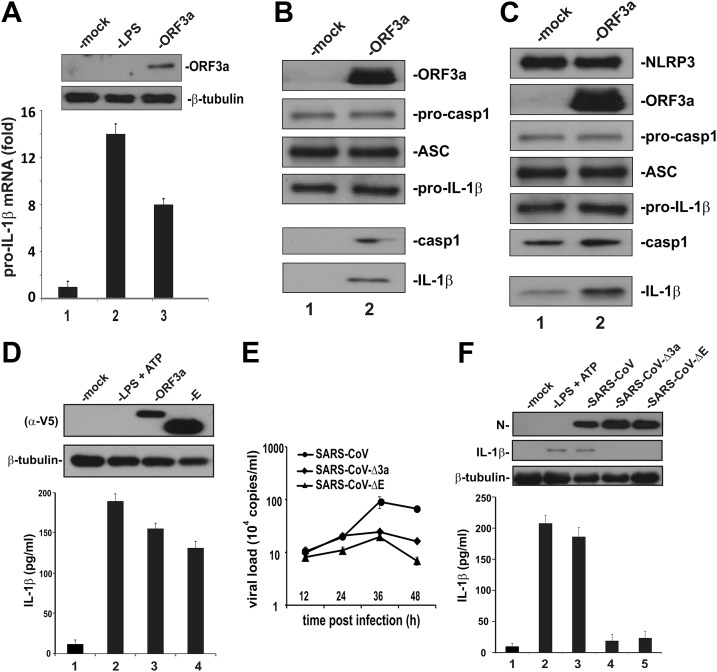
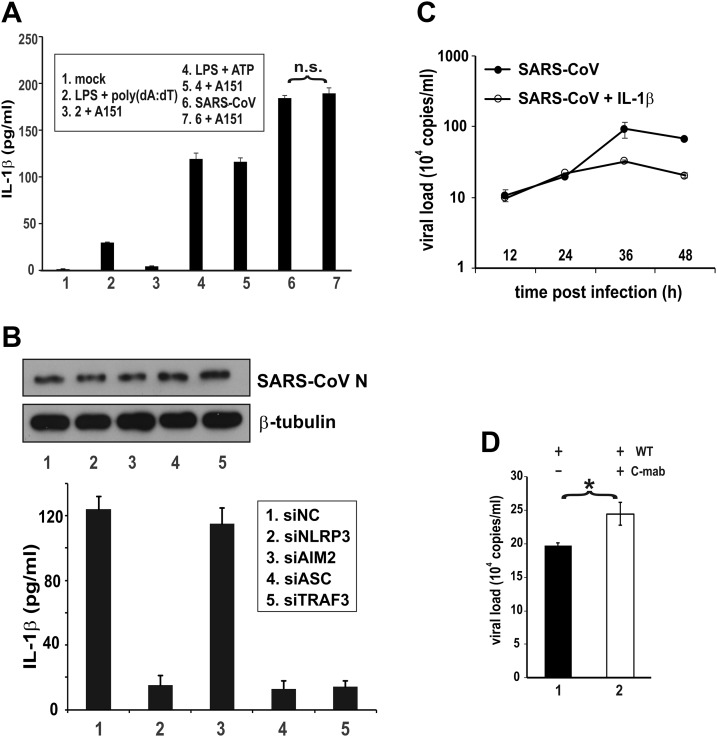
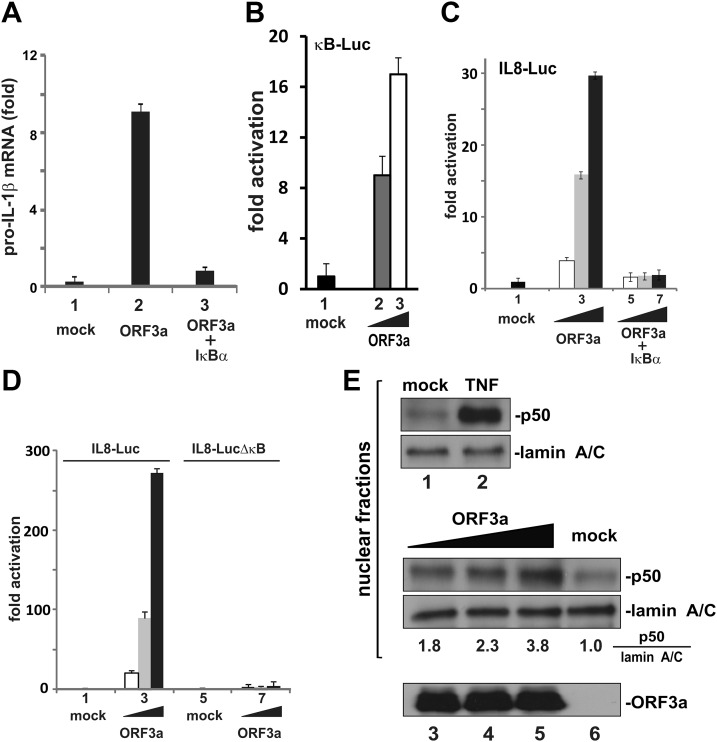
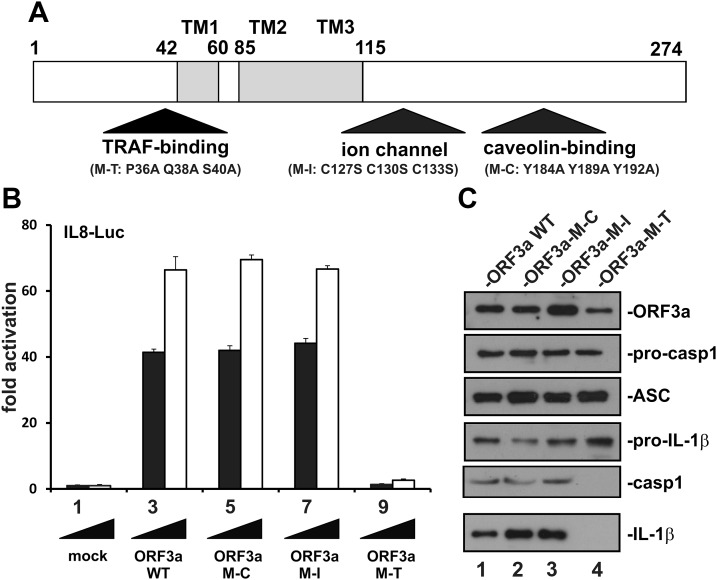
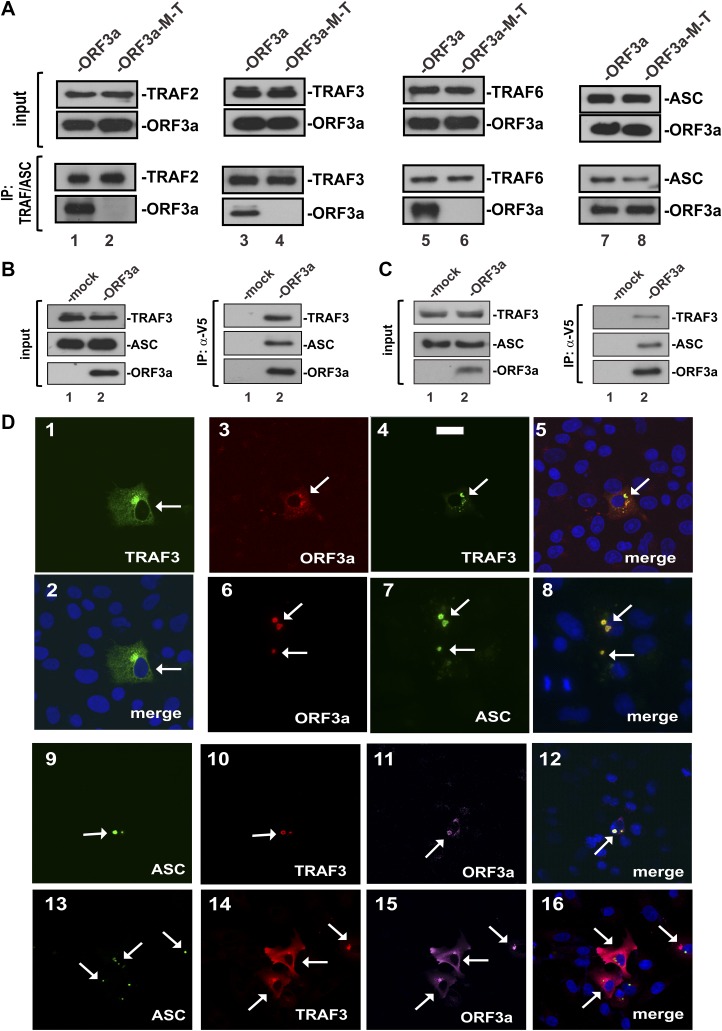
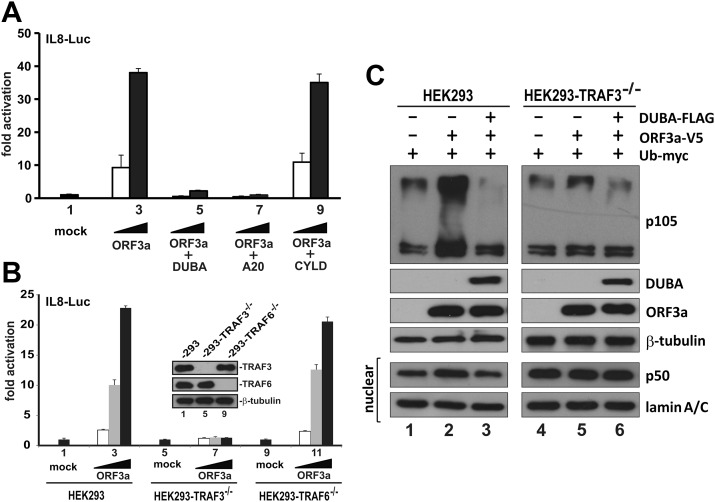
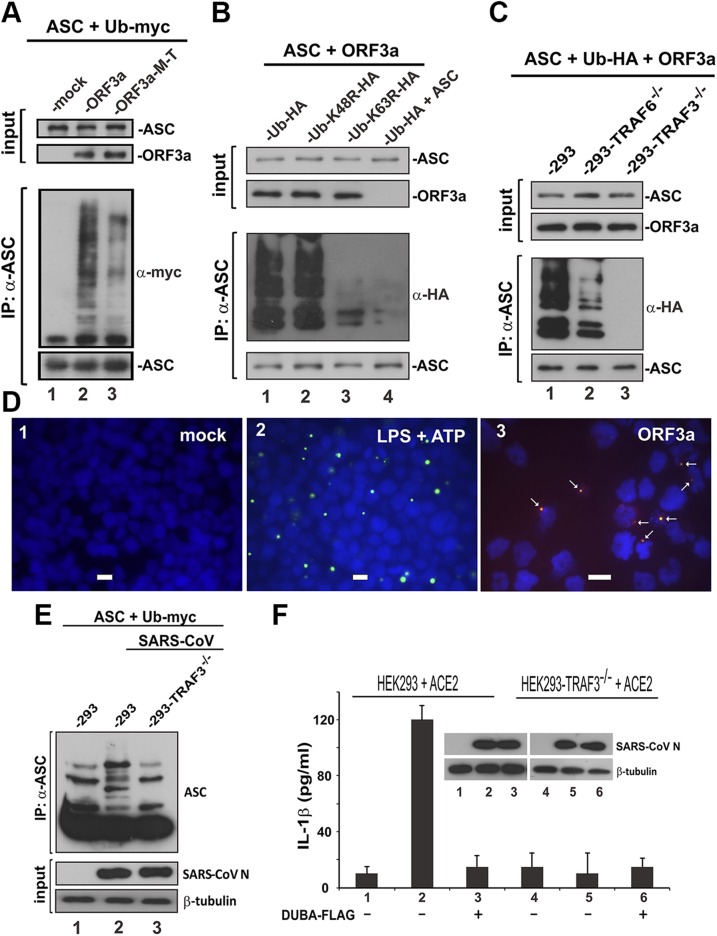
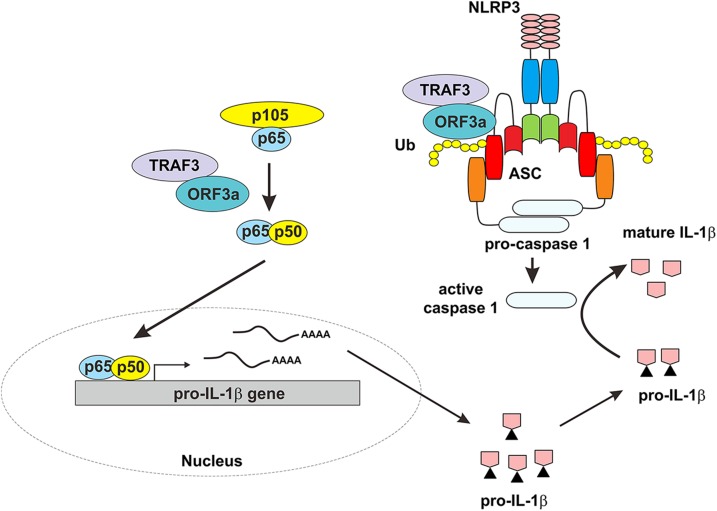
Severe acute respiratory syndrome coronavirus (SARS-CoV) is capable of inducing a storm of proinflammatory cytokines. In this study, we show that the SARS-CoV open reading frame 3a (ORF3a) accessory protein activates the NLRP3 inflammasome by promoting TNF receptor-associated factor 3 (TRAF3)-mediated ubiquitination of apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC). SARS-CoV and its ORF3a protein were found to be potent activators of pro-IL-1β gene transcription and protein maturation, the 2 signals required for activation of the NLRP3 inflammasome. ORF3a induced pro-IL-1β transcription through activation of NF-κB, which was mediated by TRAF3-dependent ubiquitination and processing of p105. ORF3a-induced elevation of IL-1β secretion was independent of its ion channel activity or absent in melanoma 2 but required NLRP3, ASC, and TRAF3. ORF3a interacted with TRAF3 and ASC, colocalized with them in discrete punctate structures in the cytoplasm, and facilitated ASC speck formation. TRAF3-dependent K63-linked ubiquitination of ASC was more pronounced in SARS-CoV-infected cells or when ORF3a was expressed. Taken together, our findings reveal a new mechanism by which SARS-CoV ORF3a protein activates NF-κB and the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of p105 and ASC.-Siu, K.-L., Yuen, K.-S., Castaño-Rodriguez, C., Ye, Z.-W., Yeung, M.-L., Fung, S.-Y., Yuan, S., Chan, C.-P., Yuen, K.-Y., Enjuanes, L., Jin, D.-Y. Severe acute respiratory syndrome coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC.
Keywords: SARS coronavirus.
Conflict of interest statement
The authors thank Lilong Jia (Department of Microbiology, The University of Hong Kong) for assistance with the infection study. This work was supported by the Hong Kong Health and Medical Research Fund (13121032, 14130822, and HKM-15-M01), Hong Kong Research Grants Council (T11-707/15-R, C7011-15R, and 17124415), Government of Spain (BIO2013-42869-R), U.S. National Institutes of Health (NIH) National Institute of Allergy and Infectious Diseases (2P01AI060699-06A1), and European Commission (Innovative Medicines Initiative Grant Agreement 115760). C.C.-R. received a fellowship from Fundacion La Caixa. K.-L.S., K.-S.Y., and C.C.-R. are co-first authors. The authors declare no conflicts of interest.
Figures
References
-
- Tang B. S., Chan K. H., Cheng V. C., Woo P. C., Lau S. K., Lam C. C., Chan T. L., Wu A. K., Hung I. F., Leung S. Y., Yuen K. Y. (2005) Comparative host gene transcription by microarray analysis early after infection of the Huh7 cell line by severe acute respiratory syndrome coronavirus and human coronavirus 229E. J. Virol. 79, 6180–6193 - PMC - PubMed
-
- Broz P., Dixit V. M. (2016) Inflammasomes: mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 16, 407–420 - PubMed
-
- Agostini L., Martinon F., Burns K., McDermott M. F., Hawkins P. N., Tschopp J. (2004) NALP3 forms an IL-1β-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity 20, 319–325 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
