

The opposing effects of interferon-beta and oncostatin-M as regulators of cancer stem cell plasticity in triple-negative breast cancer
- PMID: 31036052
- PMCID: PMC6489282
- DOI: 10.1186/s13058-019-1136-x
The opposing effects of interferon-beta and oncostatin-M as regulators of cancer stem cell plasticity in triple-negative breast cancer
Abstract
Background: Highly aggressive, metastatic and therapeutically resistant triple-negative breast cancers (TNBCs) are often enriched for cancer stem cells (CSC). Cytokines within the breast tumor microenvironment (TME) influence the CSC state by regulating tumor cell differentiation programs. Two prevalent breast TME cytokines are oncostatin-M (OSM) and interferon-β (IFN-β). OSM is a member of the IL-6 family of cytokines and can drive the de-differentiation of TNBC cells to a highly aggressive CSC state. Conversely, IFN-β induces the differentiation of TNBC, resulting in the repression of CSC properties. Here, we assess how these breast TME cytokines influence CSC plasticity and clinical outcome.
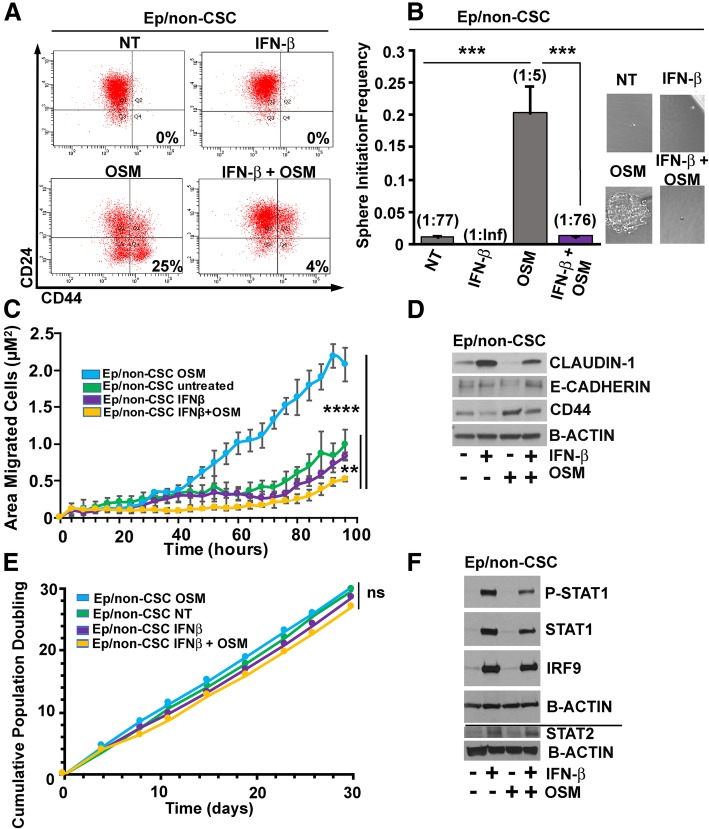
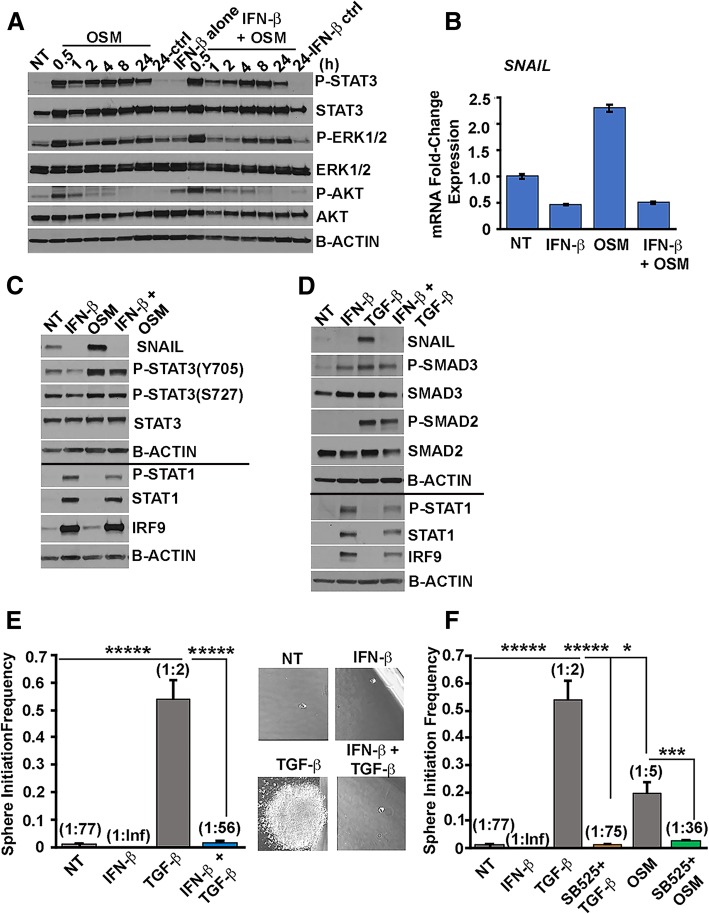
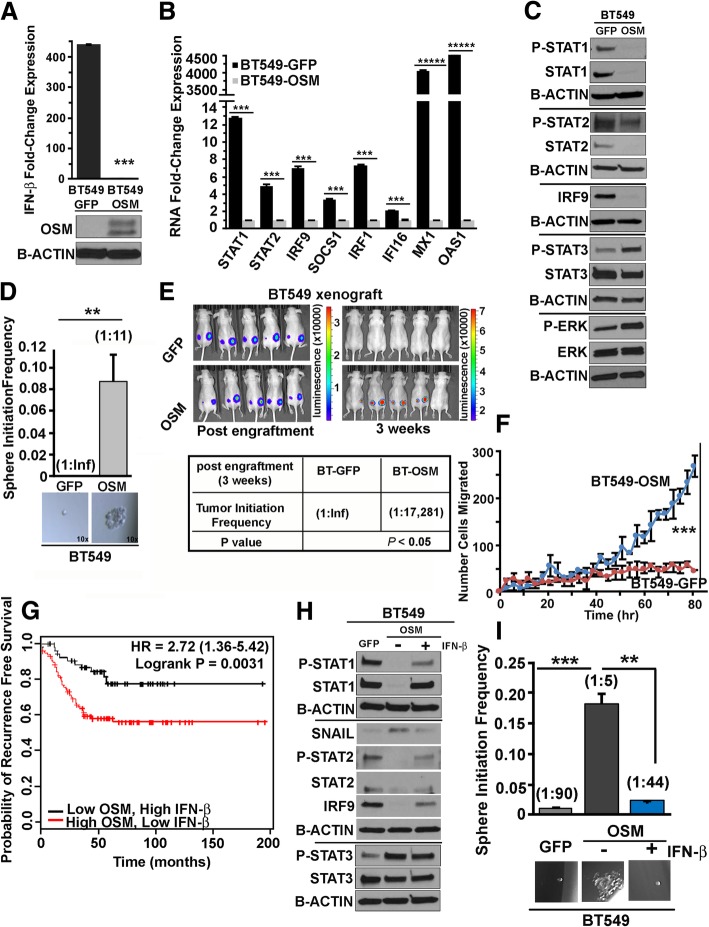
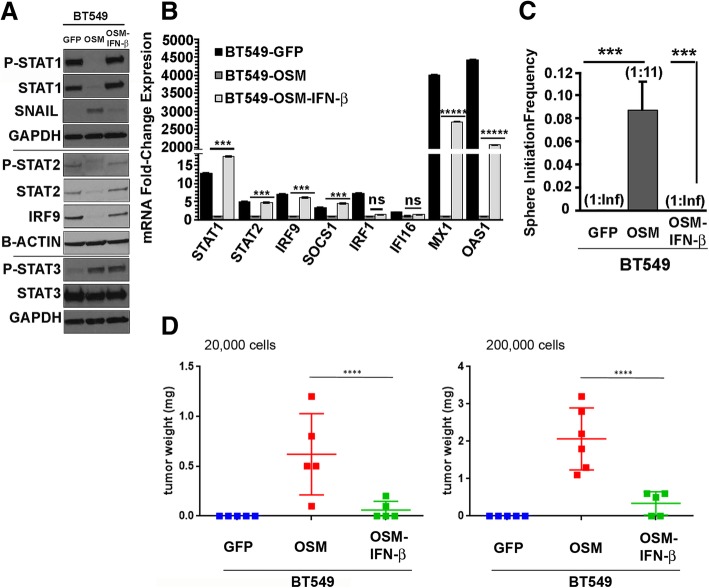
Methods: Using transformed human mammary epithelial cell (HMEC) and TNBC cell models, we assessed the CSC markers and properties following exposure to OSM and/or IFN-β. CSC markers included CD24, CD44, and SNAIL; CSC properties included tumor sphere formation, migratory capacity, and tumor initiation.
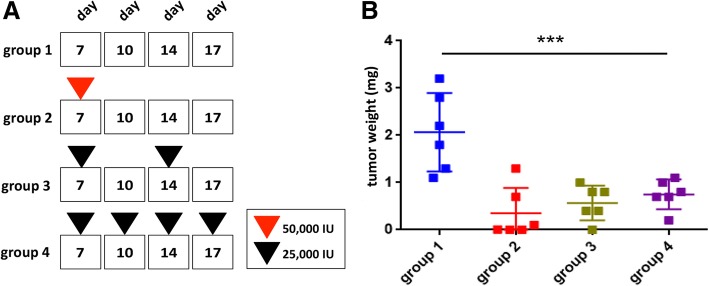
Results: There are three major findings from our study. First, exposure of purified, non-CSC to IFN-β prevents OSM-mediated CD44 and SNAIL expression and represses tumor sphere formation and migratory capacity. Second, during OSM-induced de-differentiation, OSM represses endogenous IFN-β mRNA expression and autocrine/paracrine IFN-β signaling. Restoring IFN-β signaling to OSM-driven CSC re-engages IFN-β-mediated differentiation by repressing OSM/STAT3/SMAD3-mediated SNAIL expression, tumor initiation, and growth. Finally, the therapeutic use of IFN-β to treat OSM-driven tumors significantly suppresses tumor growth.
Conclusions: Our findings suggest that the levels of IFN-β and OSM in TNBC dictate the abundance of cells with a CSC phenotype. Indeed, TNBCs with elevated IFN-β signaling have repressed CSC properties and a better clinical outcome. Conversely, TNBCs with elevated OSM signaling have a worse clinical outcome. Likewise, since OSM suppresses IFN-β expression and signaling, our studies suggest that strategies to limit OSM signaling or activate IFN-β signaling will disengage the de-differentiation programs responsible for the aggressiveness of TNBCs.
Keywords: Cancer stem cell plasticity; Cancer stem cells; Interferon-beta; Oncostatin-M; SNAIL; Triple-negative breast cancer; Tumor microenvironment.
Conflict of interest statement
Ethics approval and consent to participate
Experiments involving animals were performed in compliance with the guidelines approved by the Case Western Reserve University Institutional Animal Care and Use Committee (IACUC).
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Toll-Like receptor 3-mediated interferon-β production is suppressed by oncostatin m and a broader epithelial-mesenchymal transition program.Breast Cancer Res. 2024 Nov 26;26(1):167. doi: 10.1186/s13058-024-01918-2. Breast Cancer Res. 2024. PMID: 39593161 Free PMC article.
-
Interferon-beta represses cancer stem cell properties in triple-negative breast cancer.Proc Natl Acad Sci U S A. 2017 Dec 26;114(52):13792-13797. doi: 10.1073/pnas.1713728114. Epub 2017 Dec 11. Proc Natl Acad Sci U S A. 2017. PMID: 29229854 Free PMC article.
-
Potent EMT and CSC Phenotypes Are Induced By Oncostatin-M in Pancreatic Cancer.Mol Cancer Res. 2017 Apr;15(4):478-488. doi: 10.1158/1541-7786.MCR-16-0337. Epub 2017 Jan 4. Mol Cancer Res. 2017. PMID: 28053127 Free PMC article.
-
Developmental signaling pathways regulating mammary stem cells and contributing to the etiology of triple-negative breast cancer.Breast Cancer Res Treat. 2016 Apr;156(2):211-26. doi: 10.1007/s10549-016-3746-7. Epub 2016 Mar 11. Breast Cancer Res Treat. 2016. PMID: 26968398 Free PMC article. Review.
-
Oncostatin M: A mysterious cytokine in cancers.Int Immunopharmacol. 2021 Jan;90:107158. doi: 10.1016/j.intimp.2020.107158. Epub 2020 Nov 10. Int Immunopharmacol. 2021. PMID: 33187910 Review.
Cited by
-
Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies.Front Immunol. 2020 Aug 7;11:1280. doi: 10.3389/fimmu.2020.01280. eCollection 2020. Front Immunol. 2020. PMID: 32849491 Free PMC article. Review.
-
Dissecting the emerging role of cancer-associated adipocyte-derived cytokines in remodeling breast cancer progression.Heliyon. 2024 Jul 26;10(15):e35200. doi: 10.1016/j.heliyon.2024.e35200. eCollection 2024 Aug 15. Heliyon. 2024. PMID: 39161825 Free PMC article. Review.
-
Targeting Cancer Stem Cells through Epigenetic Modulation of Interferon Response.J Pers Med. 2022 Apr 1;12(4):556. doi: 10.3390/jpm12040556. J Pers Med. 2022. PMID: 35455671 Free PMC article. Review.
-
Changes in the mammary gland during aging and its links with breast diseases.Acta Biochim Biophys Sin (Shanghai). 2023 May 15;55(6):1001-1019. doi: 10.3724/abbs.2023073. Acta Biochim Biophys Sin (Shanghai). 2023. PMID: 37184281 Free PMC article. Review.
-
How cancer cells make and respond to interferon-I.Trends Cancer. 2023 Jan;9(1):83-92. doi: 10.1016/j.trecan.2022.09.003. Epub 2022 Oct 8. Trends Cancer. 2023. PMID: 36216730 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
