

Notch signalling defines dorsal root ganglia neuroglial fate choice during early neural crest cell migration
- PMID: 31036074
- PMCID: PMC6489353
- DOI: 10.1186/s12868-019-0501-0
Notch signalling defines dorsal root ganglia neuroglial fate choice during early neural crest cell migration
Abstract
Background: The dorsal root ganglia (DRG) are a critical component of the peripheral nervous system, and function to relay somatosensory information from the body's periphery to sensory perception centres within the brain. The DRG are primarily comprised of two cell types, sensory neurons and glia, both of which are neural crest-derived. Notch signalling is known to play an essential role in defining the neuronal or glial fate of bipotent neural crest progenitors that migrate from the dorsal ridge of the neural tube to the sites of the DRG. However, the involvement of Notch ligands in this process and the timing at which neuronal versus glial fate is acquired has remained uncertain.
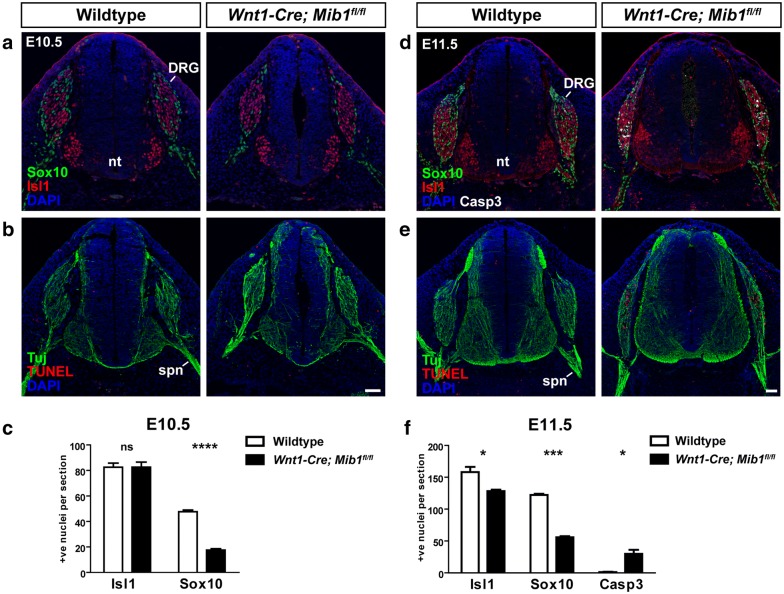
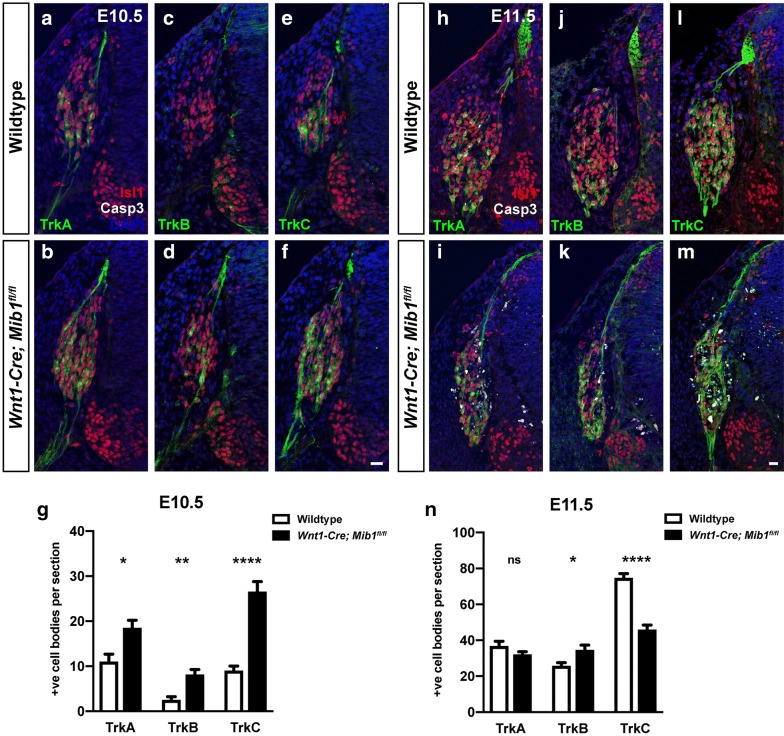
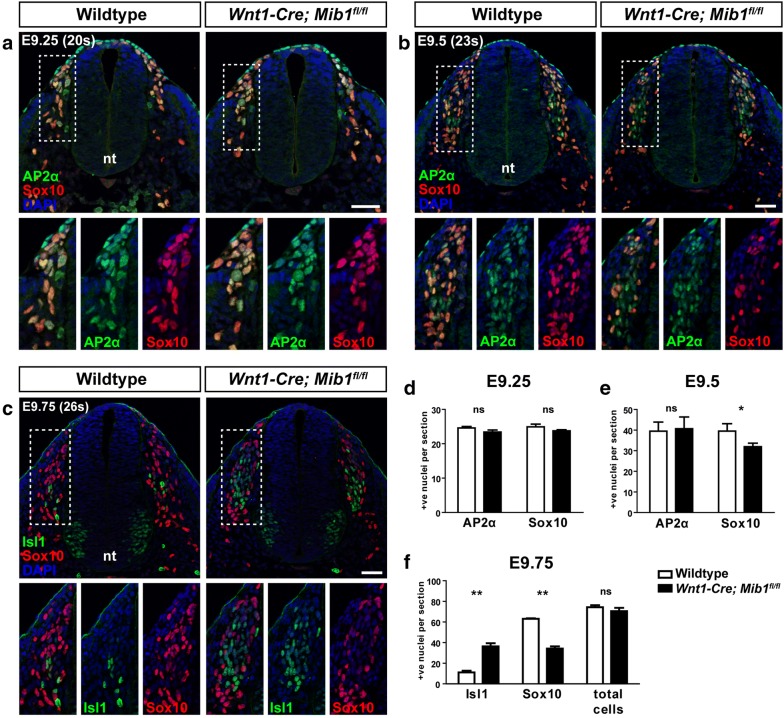
Results: We have used tissue specific knockout of the E3 ubiquitin ligase mindbomb1 (Mib1) to remove the function of all Notch ligands in neural crest cells. Wnt1-Cre; Mib1fl/fl mice exhibit severe DRG defects, including a reduction in glial cells, and neuronal cell death later in development. By comparing formation of sensory neurons and glia with the expression and activation of Notch signalling in these mice, we define a critical period during embryonic development in which early migrating neural crest cells become biased toward neuronal and glial phenotypes.
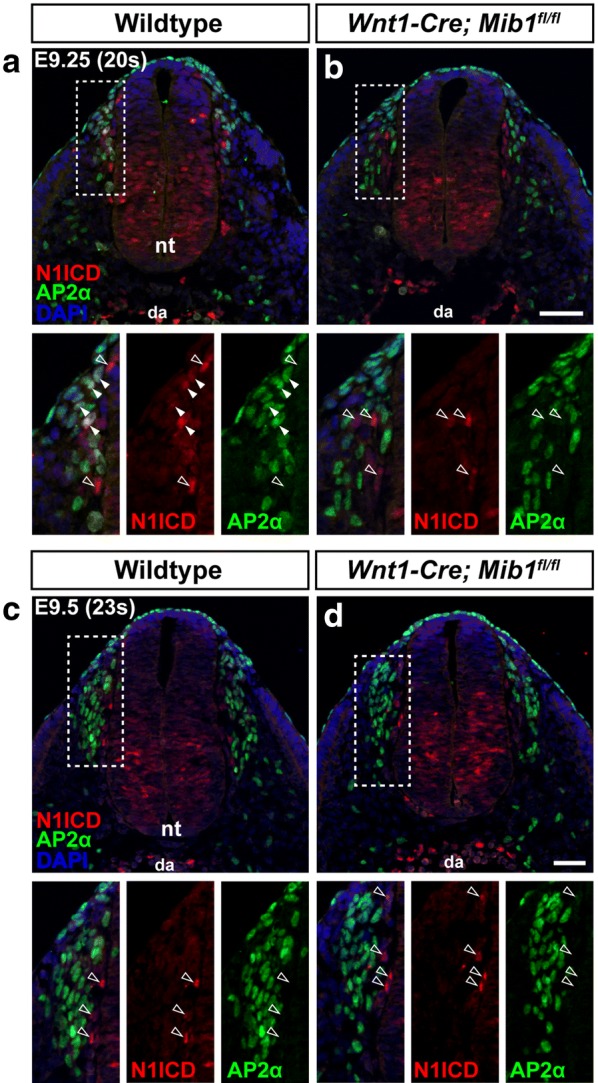
Conclusions: We demonstrate active Notch signalling between neural crest progenitors as soon as trunk neural crest cells delaminate from the neural tube and during their early migration toward the site of the DRG. This data brings into question the timing of neuroglial fate specification in the DRG and suggest that it may occur much earlier than originally considered.
Keywords: Dorsal root ganglia; Fate restriction; Mib1; Neural crest; Notch signalling.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
