

Cortical circuit alterations precede motor impairments in Huntington's disease mice
- PMID: 31036840
- PMCID: PMC6488584
- DOI: 10.1038/s41598-019-43024-w
Cortical circuit alterations precede motor impairments in Huntington's disease mice
Abstract
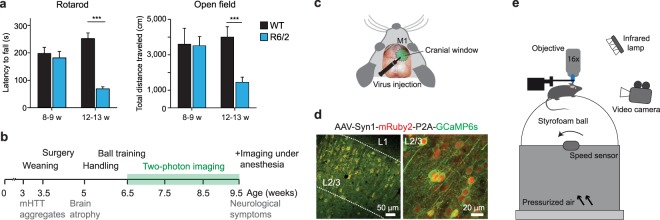
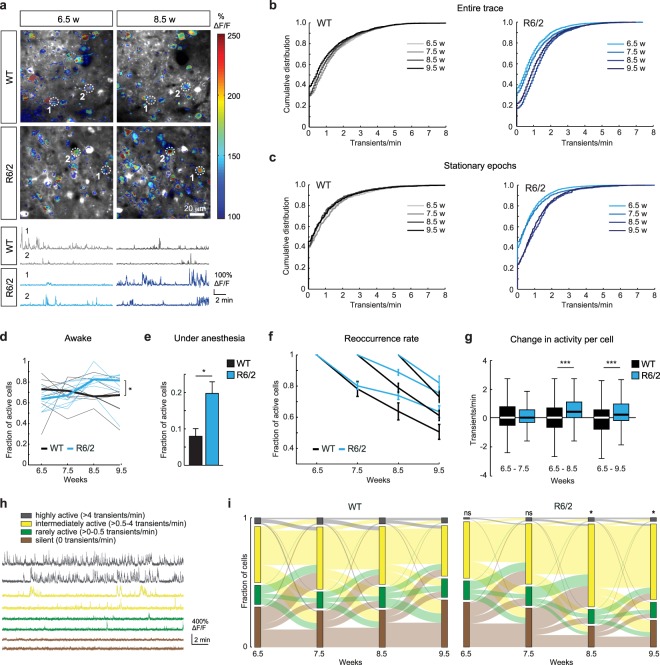
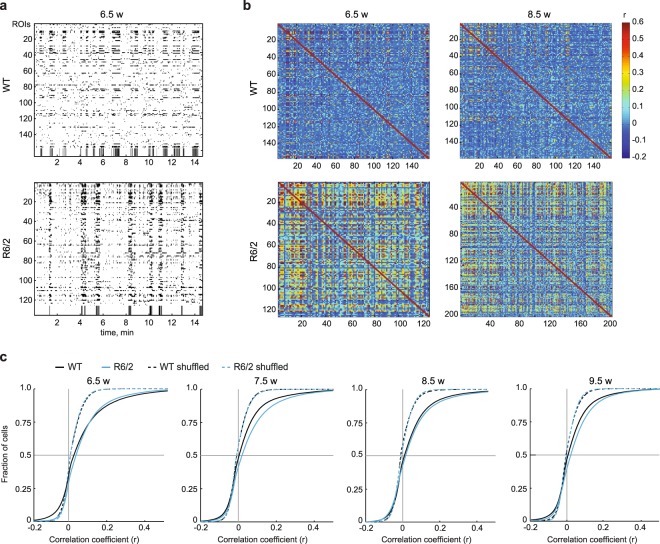
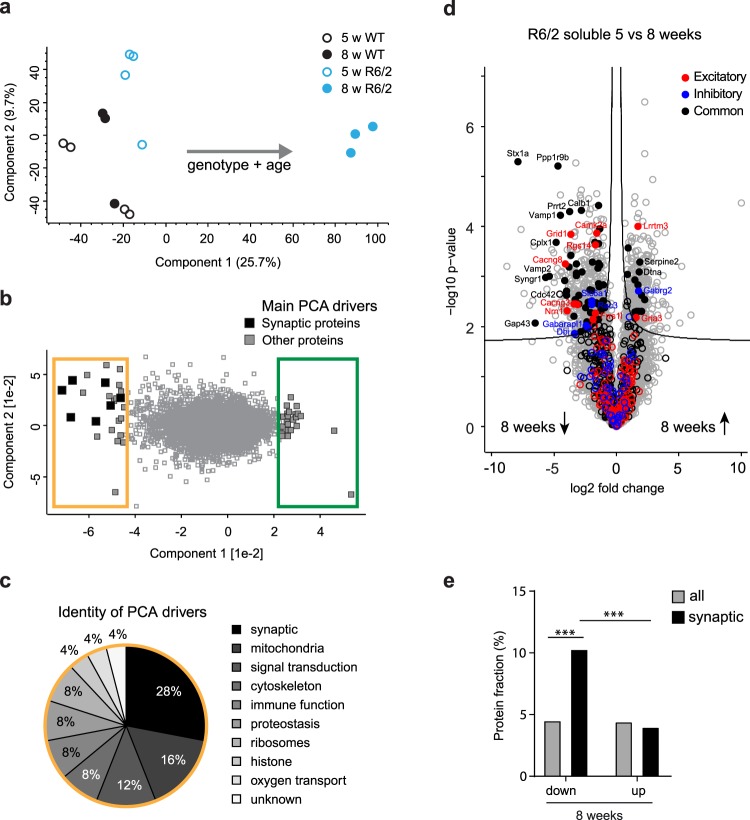
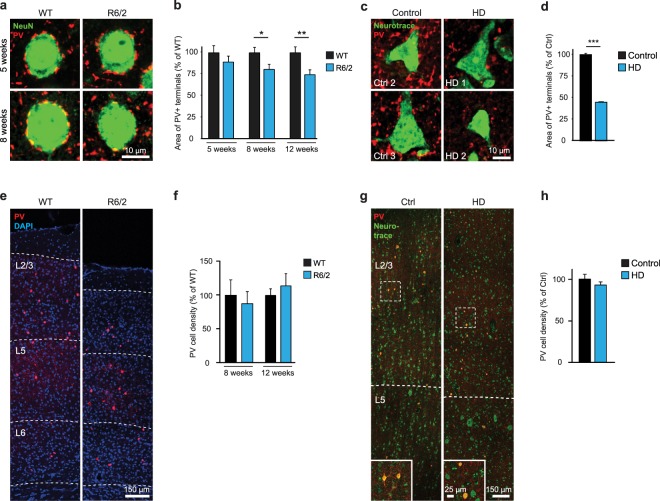
Huntington's disease (HD) is a devastating hereditary movement disorder, characterized by degeneration of neurons in the striatum and cortex. Studies in human patients and mouse HD models suggest that disturbances of neuronal function in the neocortex play an important role in disease onset and progression. However, the precise nature and time course of cortical alterations in HD have remained elusive. Here, we use chronic in vivo two-photon calcium imaging to longitudinally monitor the activity of identified single neurons in layer 2/3 of the primary motor cortex in awake, behaving R6/2 transgenic HD mice and wildtype littermates. R6/2 mice show age-dependent changes in cortical network function, with an increase in activity that affects a large fraction of cells and occurs rather abruptly within one week, preceeding the onset of motor defects. Furthermore, quantitative proteomics demonstrate a pronounced downregulation of synaptic proteins in the cortex, and histological analyses in R6/2 mice and human HD autopsy cases reveal a reduction in perisomatic inhibitory synaptic contacts on layer 2/3 pyramidal cells. Taken together, our study provides a time-resolved description of cortical network dysfunction in behaving HD mice and points to disturbed excitation/inhibition balance as an important pathomechanism in HD.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
