

NF-κB signaling dynamics is controlled by a dose-sensing autoregulatory loop
- PMID: 31040261
- PMCID: PMC7083554
- DOI: 10.1126/scisignal.aau3568
NF-κB signaling dynamics is controlled by a dose-sensing autoregulatory loop
Abstract
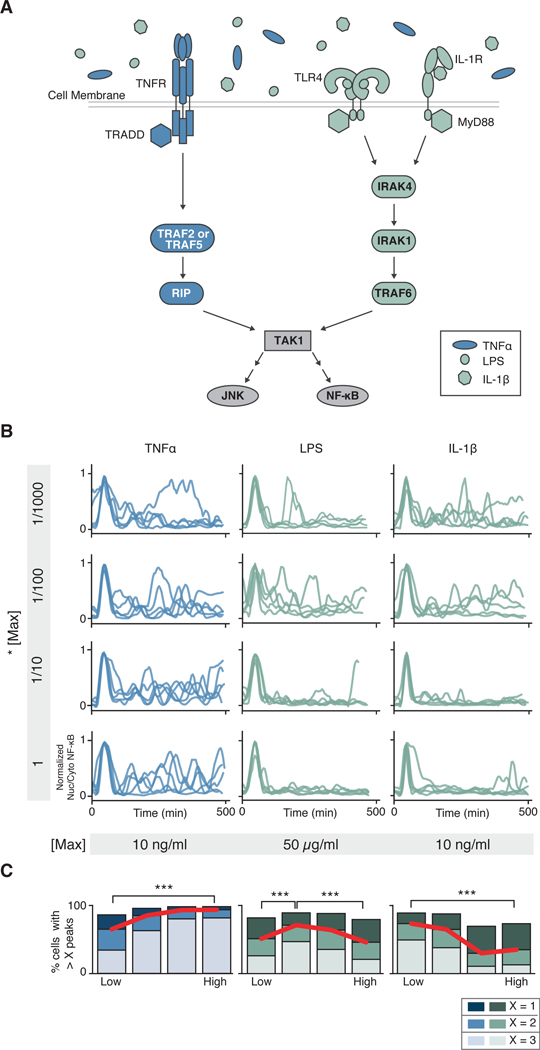
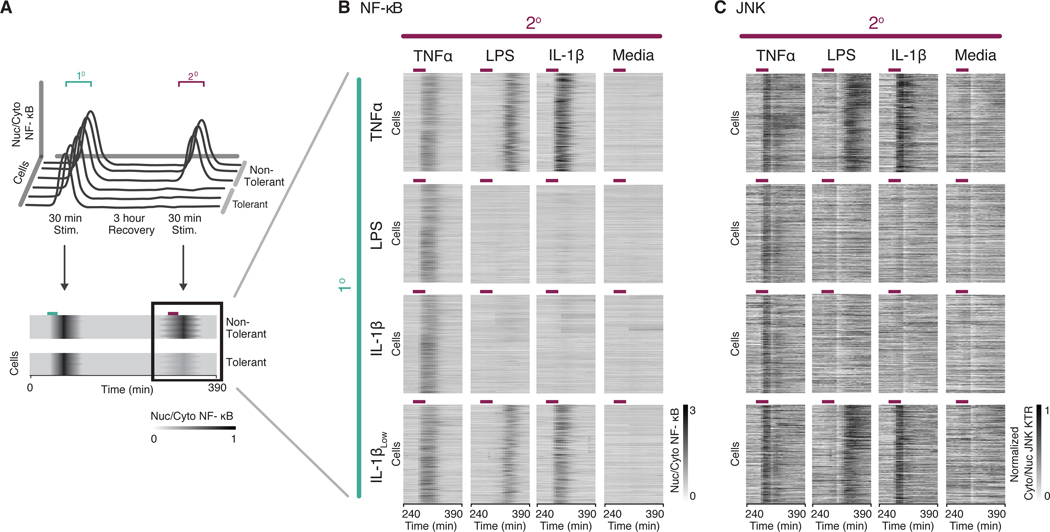
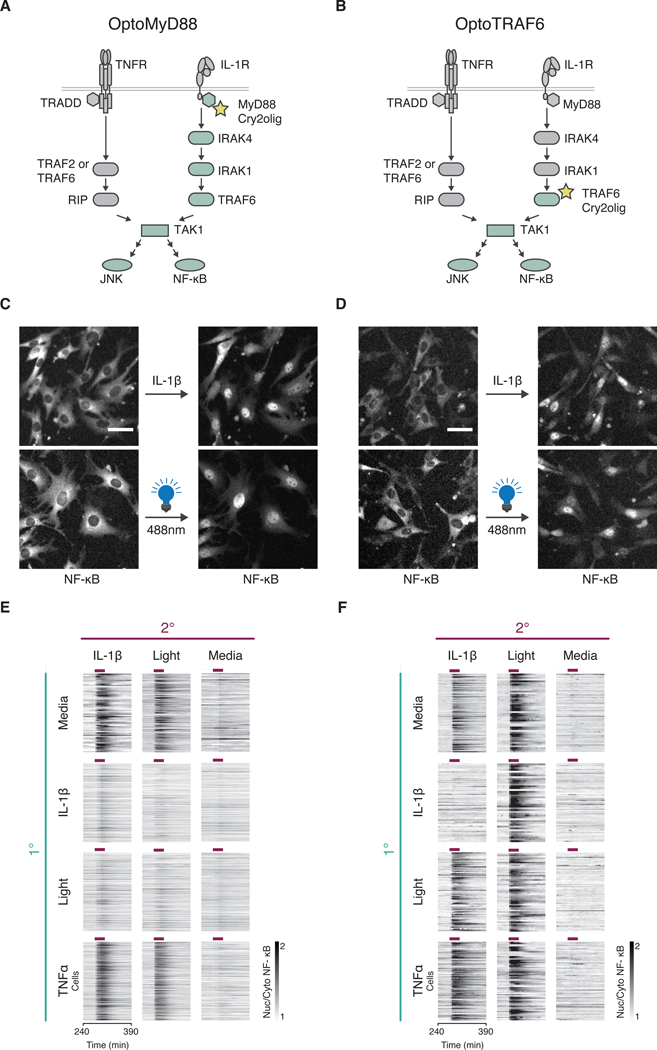
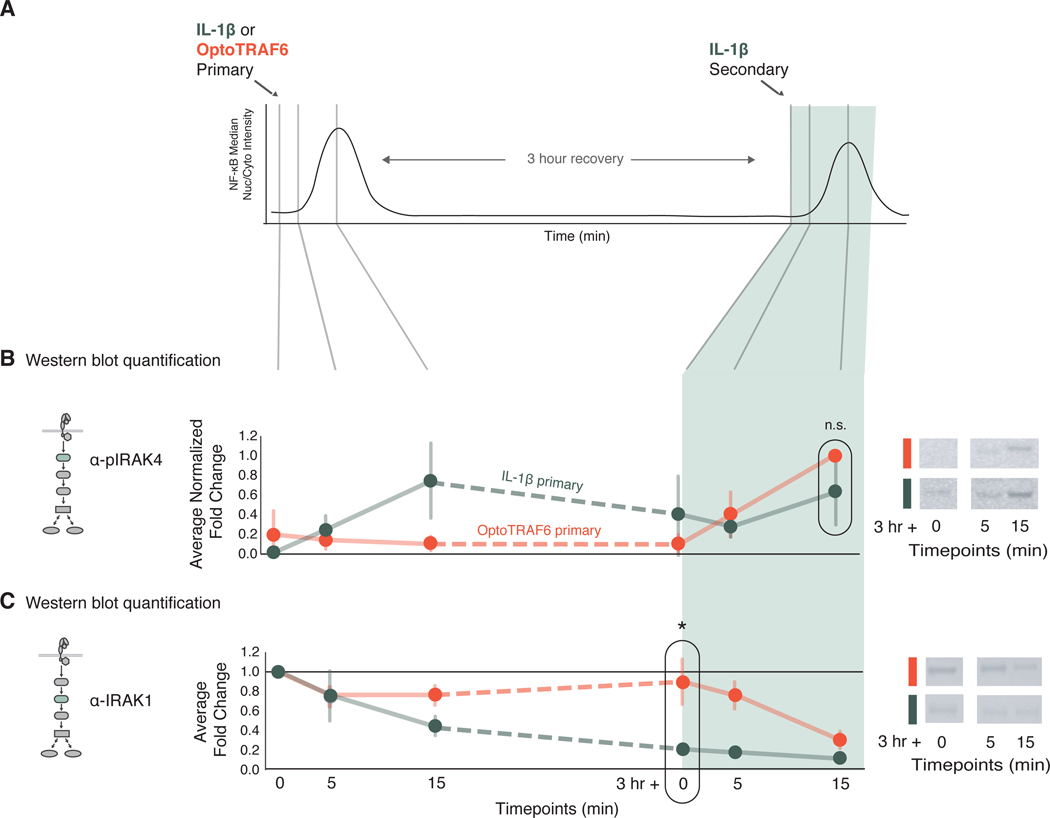
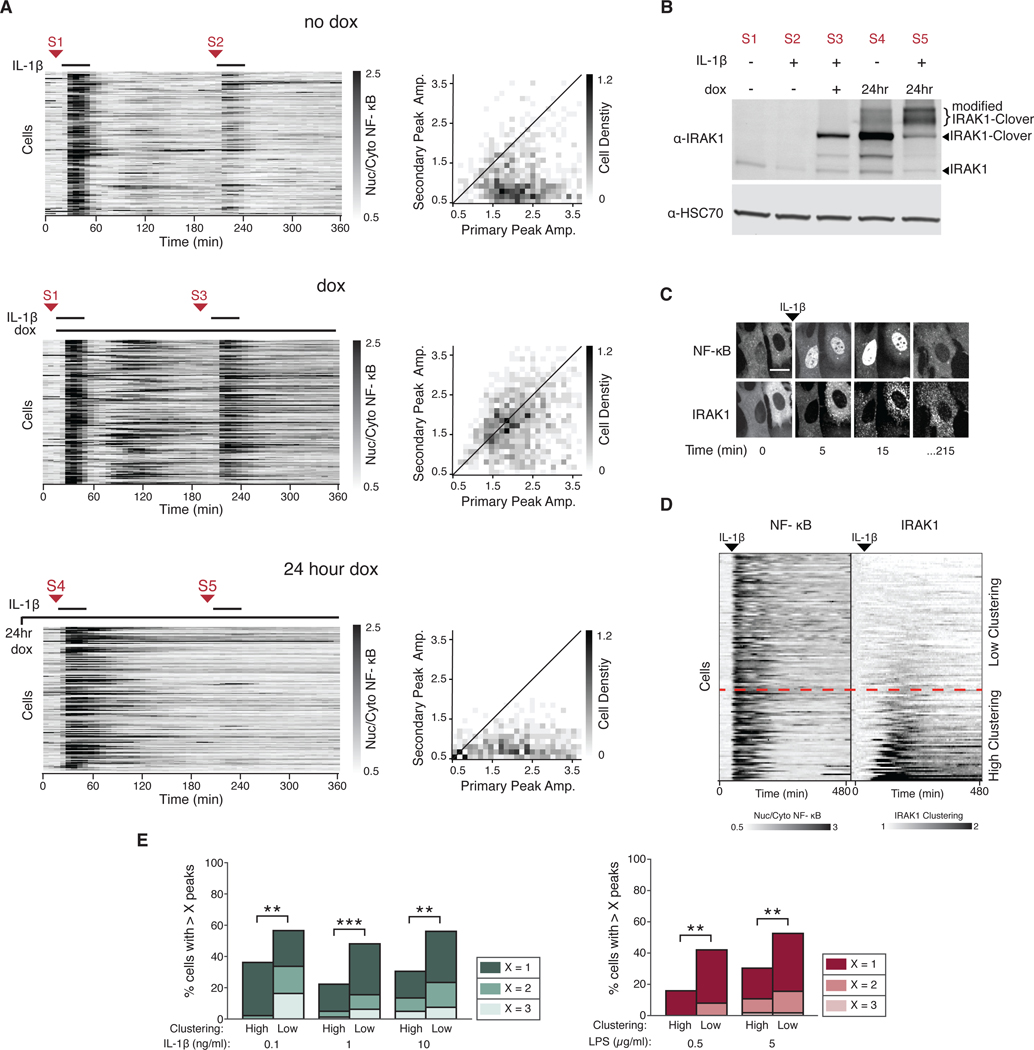
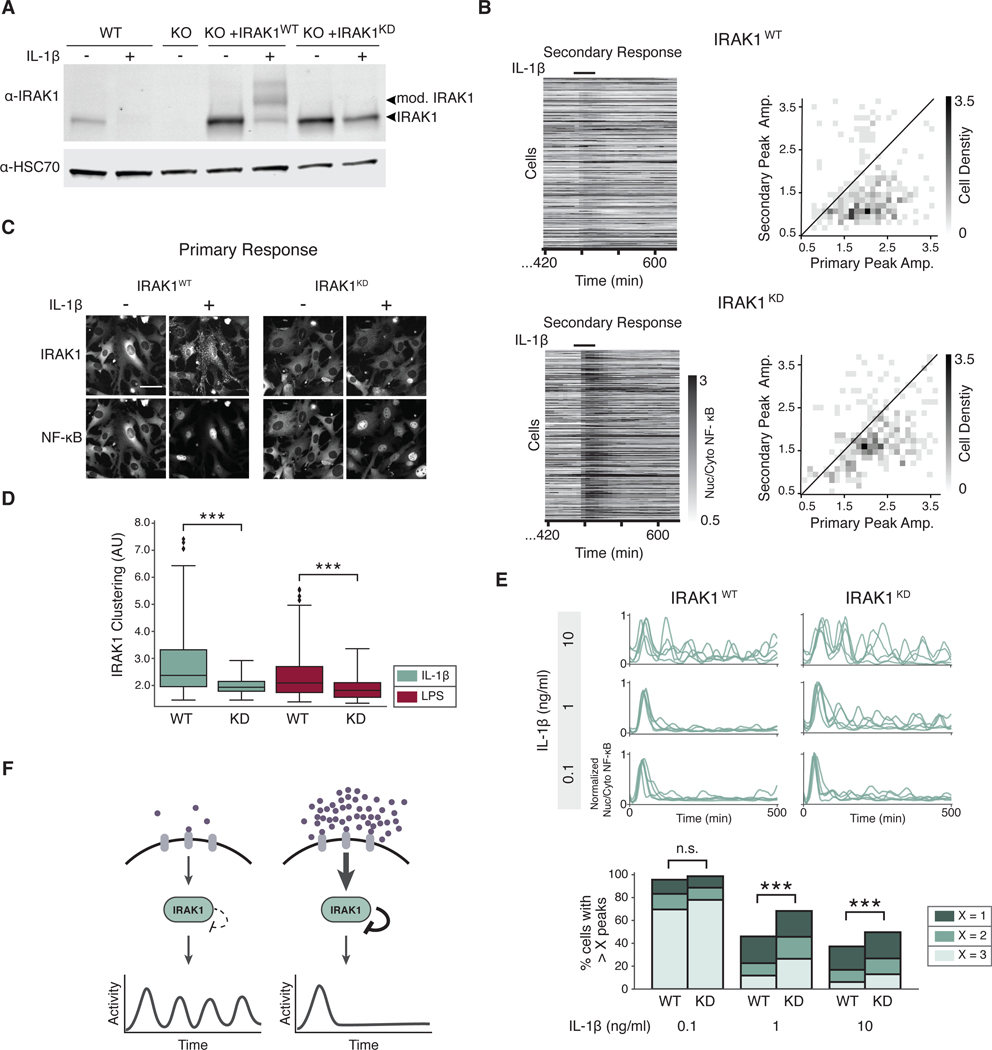
Over the last decade, multiple studies have shown that signaling proteins activated in different temporal patterns, such as oscillatory, transient, and sustained, can result in distinct gene expression patterns or cell fates. However, the molecular events that ensure appropriate stimulus- and dose-dependent dynamics are not often understood and are difficult to investigate. Here, we used single-cell analysis to dissect the mechanisms underlying the stimulus- and dose-encoding patterns in the innate immune signaling network. We found that Toll-like receptor (TLR) and interleukin-1 receptor (IL-1R) signaling dynamics relied on a dose-dependent, autoinhibitory loop that rendered cells refractory to further stimulation. Using inducible gene expression and optogenetics to perturb the network at different levels, we identified IL-1R-associated kinase 1 (IRAK1) as the dose-sensing node responsible for limiting signal flow during the innate immune response. Although the kinase activity of IRAK1 was not required for signal propagation, it played a critical role in inhibiting the nucleocytoplasmic oscillations of the transcription factor NF-κB. Thus, protein activities that may be "dispensable" from a topological perspective can nevertheless be essential in shaping the dynamic response to the external environment.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Sparta B, Pargett M, Minguet M, Distor K, Bell G, Albeck JG, Receptor Level Mechanisms Are Required for Epidermal Growth Factor (EGF)-stimulated Extracellular Signal-regulated Kinase (ERK) Activity Pulses. The Journal of biological chemistry 290, 24784–24792 (2015); published online EpubOct 09 ( 10.1074/jbc.M115.662247). - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
