

Loss of Nuclear TDP-43 Is Associated with Decondensation of LINE Retrotransposons
- PMID: 31042469
- PMCID: PMC6508629
- DOI: 10.1016/j.celrep.2019.04.003
Loss of Nuclear TDP-43 Is Associated with Decondensation of LINE Retrotransposons
Abstract
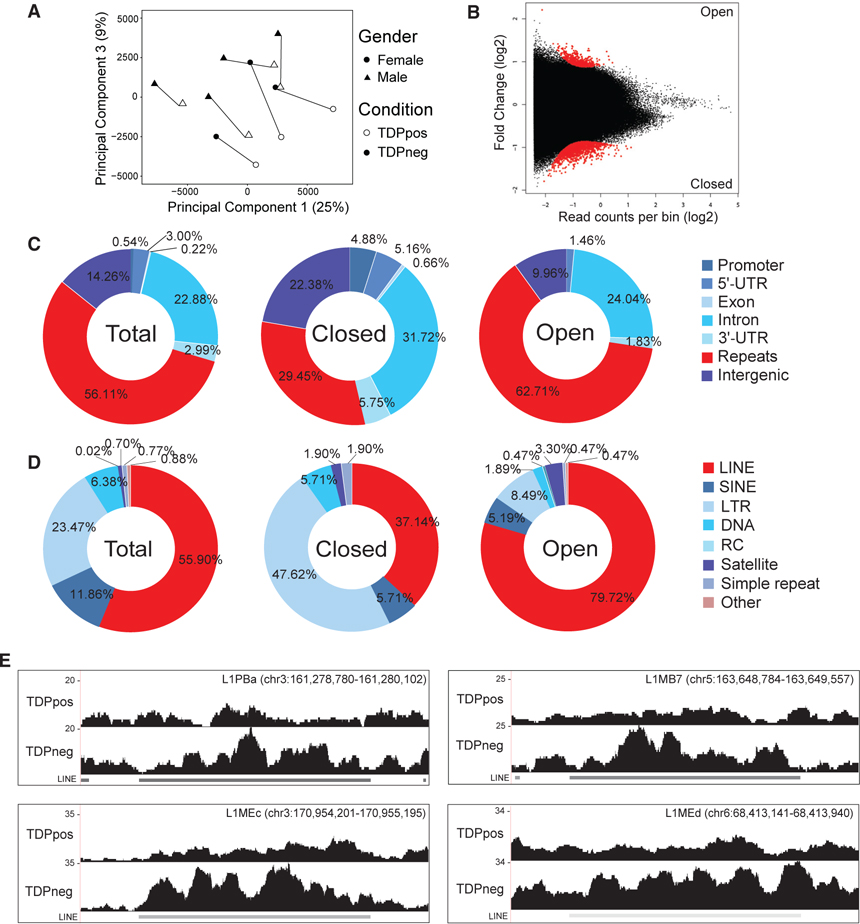
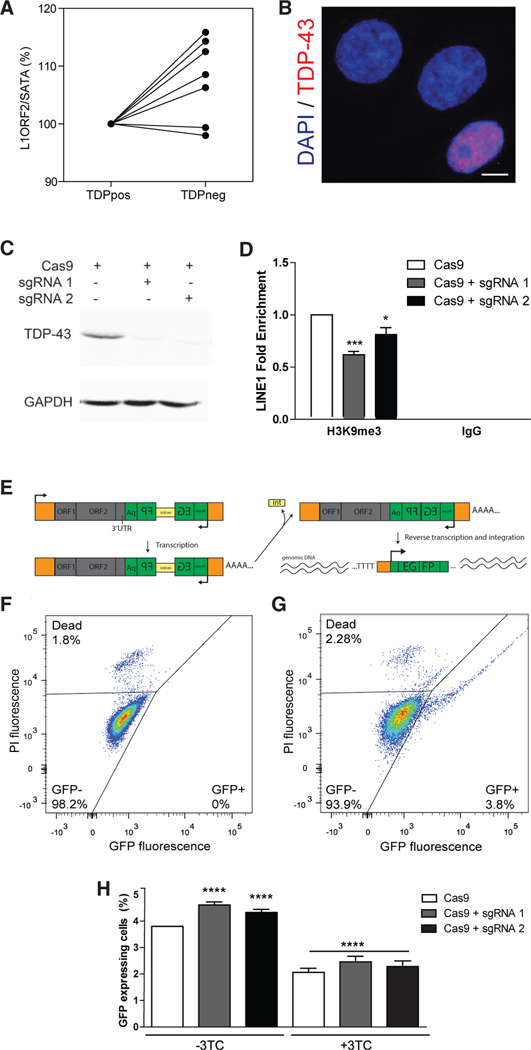
Loss of the nuclear RNA binding protein TAR DNA binding protein-43 (TDP-43) into cytoplasmic aggregates is the strongest correlate to neurodegeneration in amyotrophic lateral sclerosis and frontotemporal degeneration. The molecular changes associated with the loss of nuclear TDP-43 in human tissues are not entirely known. Using subcellular fractionation and fluorescent-activated cell sorting to enrich for diseased neuronal nuclei without TDP-43 from post-mortem frontotemporal degeneration-amyotrophic lateral sclerosis (FTD-ALS) human brain, we characterized the effects of TDP-43 loss on the transcriptome and chromatin accessibility. Nuclear TDP-43 loss is associated with gene expression changes that affect RNA processing, nucleocytoplasmic transport, histone processing, and DNA damage. Loss of nuclear TDP-43 is also associated with chromatin decondensation around long interspersed nuclear elements (LINEs) and increased LINE1 DNA content. Moreover, loss of TDP-43 leads to increased retrotransposition that can be inhibited with antiretroviral drugs, suggesting that TDP-43 neuropathology is associated with altered chromatin structure including decondensation of LINEs.
Keywords: ATAC-seq; RNA; amyotrophic lateral sclerosis; autoregulation; chromatin; frontotemporal dementia; histone; motor neuron disease; reverse transcriptase; splicing.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
E.Y.L. is currently affiliated with ETHOS Health Communications, Yardley, PA, USA. J.R. is currently affiliated with DZNE e.V., Bonn, Germany.
Figures
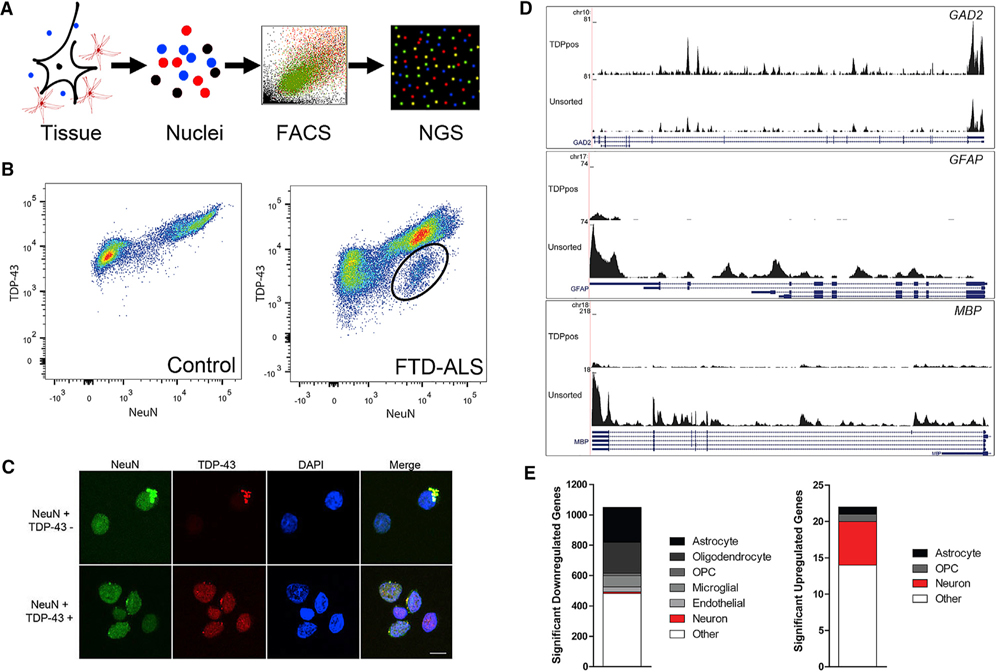
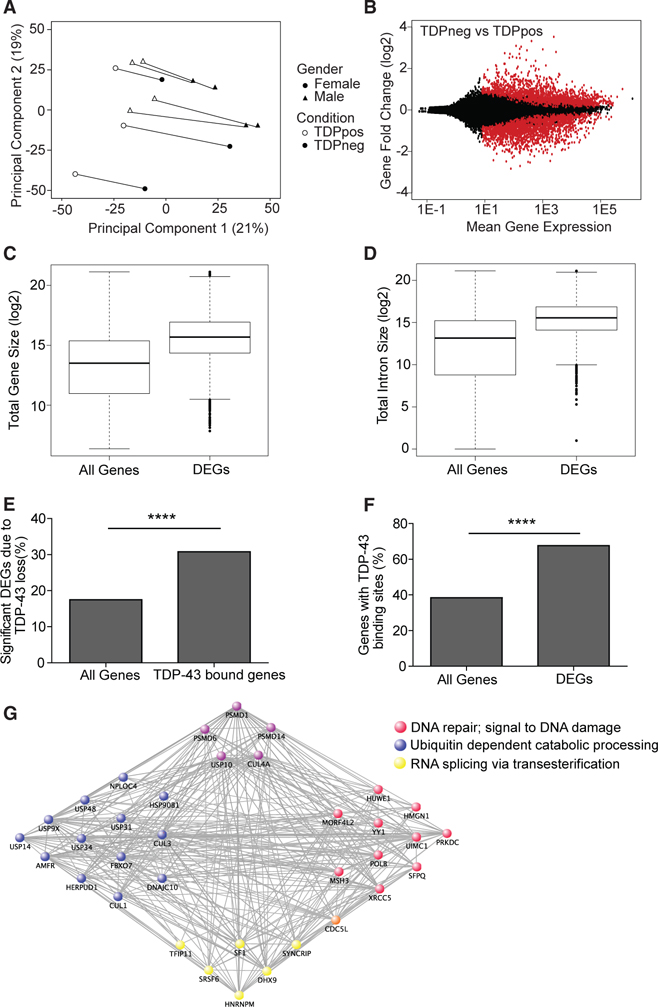
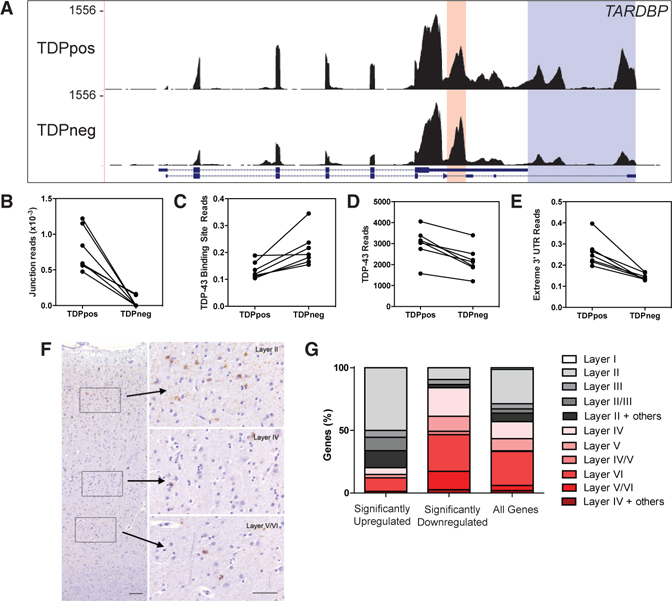
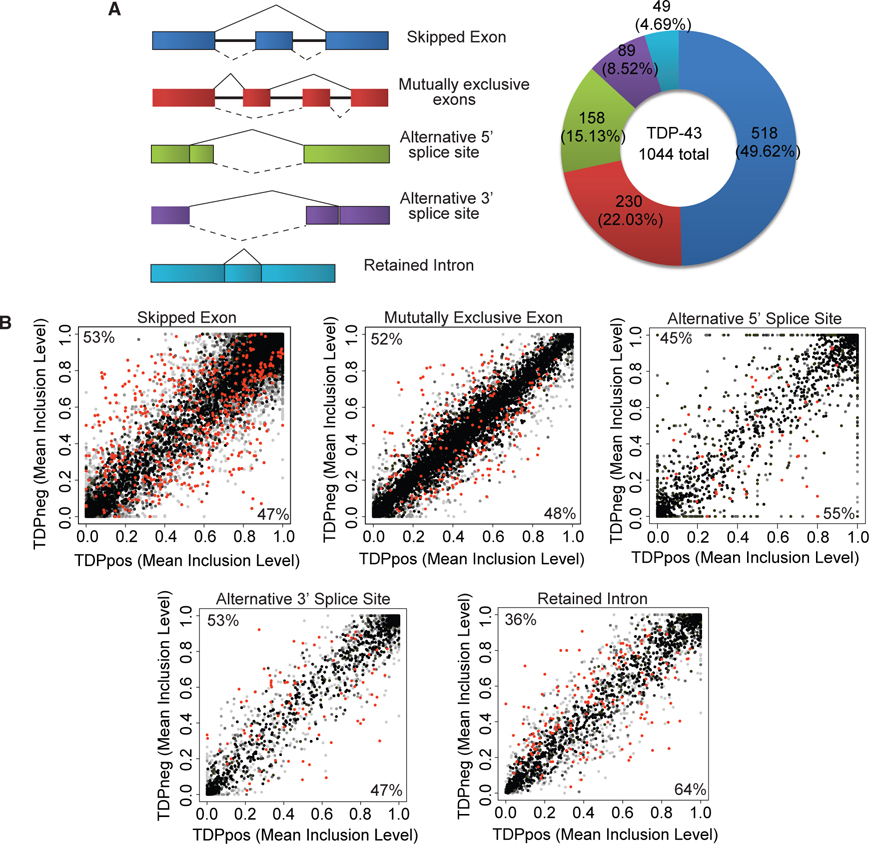
References
-
- Andrews S (2010). FastQC: a quality control tool for high throughput sequence data (Babraham Institute). http://www.bioinformatics.babraham.ac.uk/projects/fastqc.
-
- Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, Jacob Filho W, Lent R, and Herculano-Houzel S (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isomet-rically scaled-up primate brain. J. Comp. Neurol 513, 532–541. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
