

Epithelial dynamics in the epididymis: role in the maturation, protection, and storage of spermatozoa
- PMID: 31044554
- PMCID: PMC6688936
- DOI: 10.1111/andr.12632
Epithelial dynamics in the epididymis: role in the maturation, protection, and storage of spermatozoa
Abstract
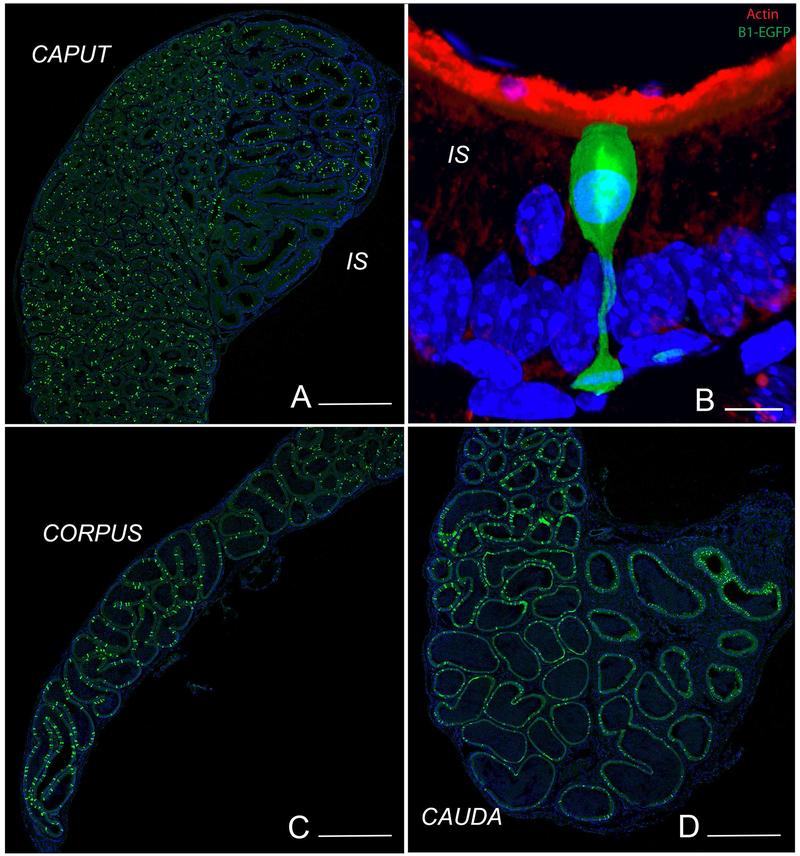
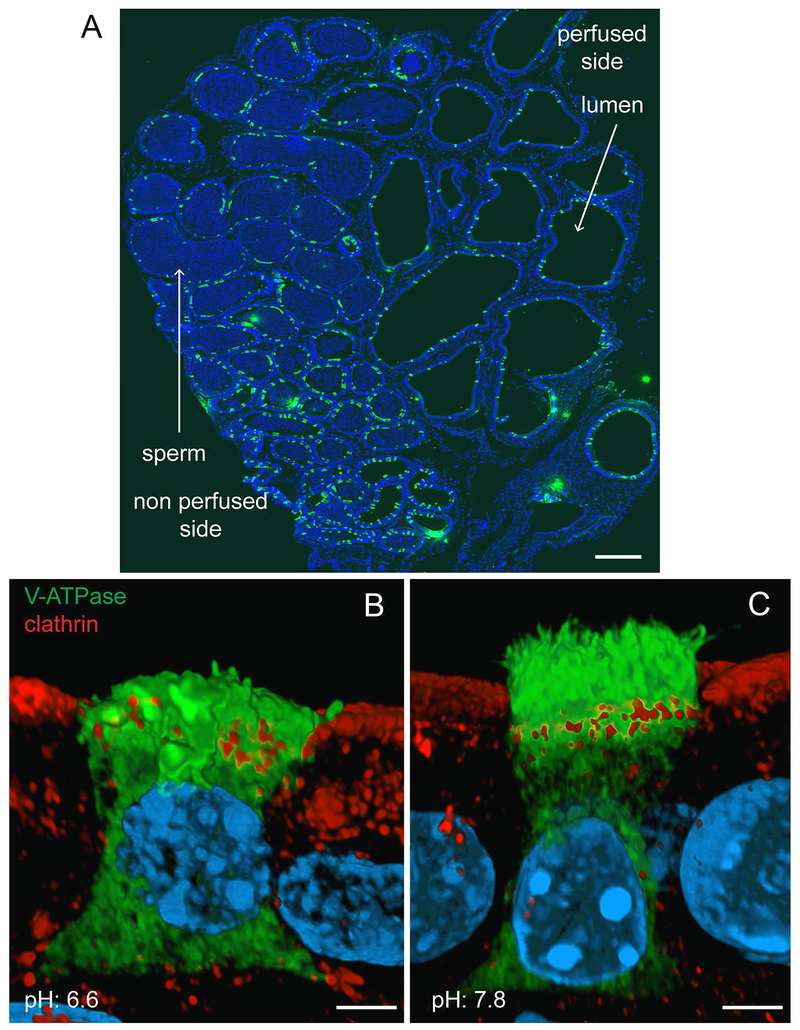
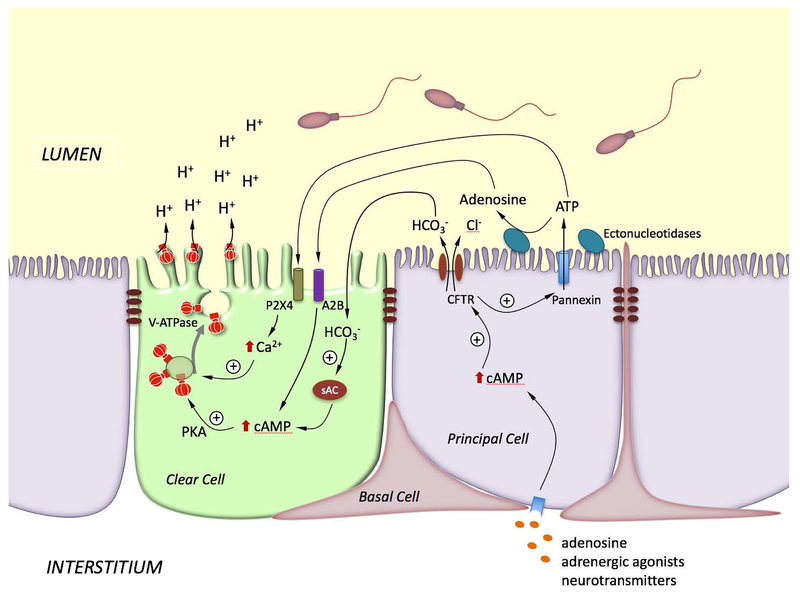
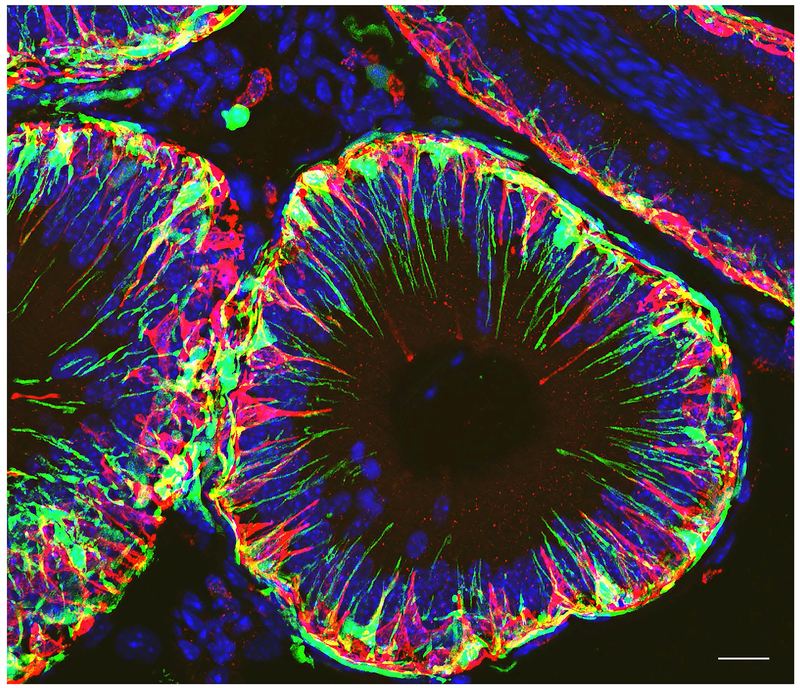
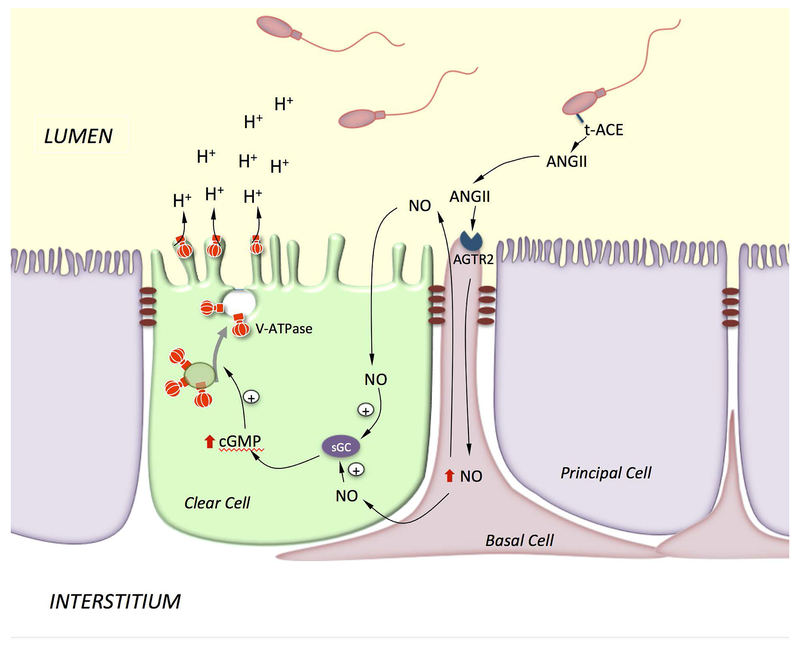
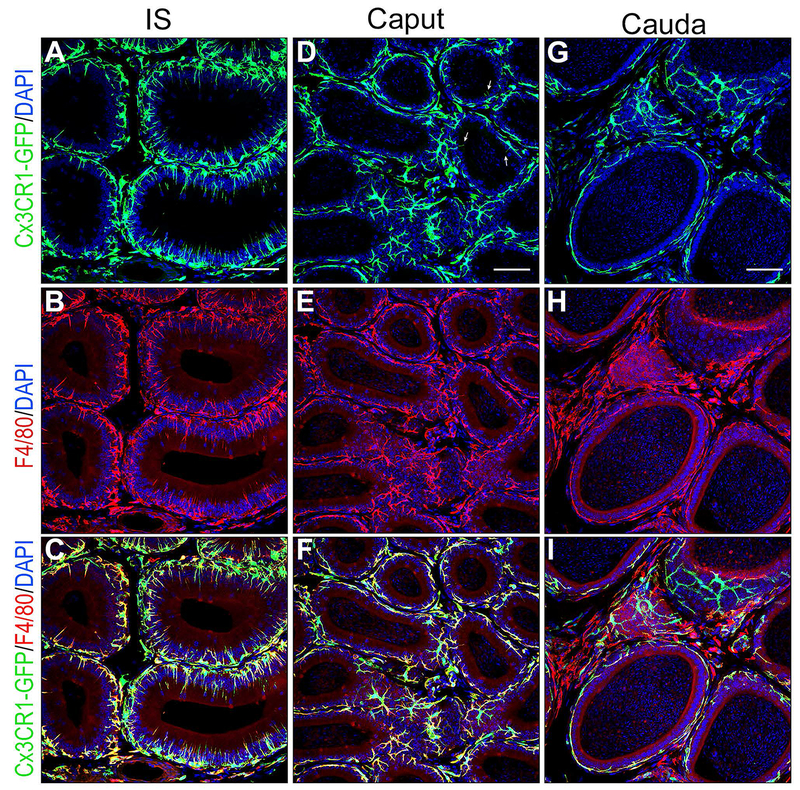
Epithelial cells line the lumen of tubular organs and are key players in their respective functions. They establish a unique luminal environment by providing a protective barrier and by performing vectorial transport of ions, nutrients, solutes, proteins, and water. Complex intercellular communication networks, specific for each organ, ensure their interaction with adjacent epithelial and non-epithelial cells, allowing them to respond to and modulate their immediate environment. In the epididymis, several epithelial cell types work in a concerted manner to establish a luminal acidic milieu that is essential for the post-testicular maturation and storage of spermatozoa. The epididymis also prevents autoimmune responses against auto-antigenic spermatozoa, while ensuring protection against ascending and blood pathogens. This is achieved by a network of immune cells that are in close contact and interact with epithelial cells. This review highlights the coordinated interactions between spermatozoa, basal cells, principal cells, narrow cells, clear cells, and immune cells that contribute to the maturation, protection, selection, and storage of spermatozoa in the lumen of the epididymis.
Keywords: epididymal immunology; luminal acidification; male fertility; post-testicular regulation; tight junctions.
© 2019 American Society of Andrology and European Academy of Andrology.
Conflict of interest statement
CONFLICT OF INTEREST
SB has a financial interest in Kantum Pharma a company developing a diagnostic and therapeutic combination to prevent and treat Acute Kidney Injury. SB’s interests were reviewed and are managed by Massachusetts General Hospital and Partners HealthCare in accordance with their conflict of interest policies.
Figures
References
-
- Bagnis C, Marsolais M, Biemesderfer D, Laprade R, Breton S. Na+/H+-exchange activity and immunolocalization of NHE3 in rat epididymis. Am J Physiol Renal Physiol 2001;280:F426–36. - PubMed
-
- Banks FC, Calvert RC, Burnstock G. Changing P2X receptor localization on maturing sperm in the epididymides of mice, hamsters, rats, and humans: a preliminary study. Fertil Steril 2010;93:1415–20. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
