

WWP2 ubiquitylates RNA polymerase II for DNA-PK-dependent transcription arrest and repair at DNA breaks
- PMID: 31048545
- PMCID: PMC6546063
- DOI: 10.1101/gad.321943.118
WWP2 ubiquitylates RNA polymerase II for DNA-PK-dependent transcription arrest and repair at DNA breaks
Abstract
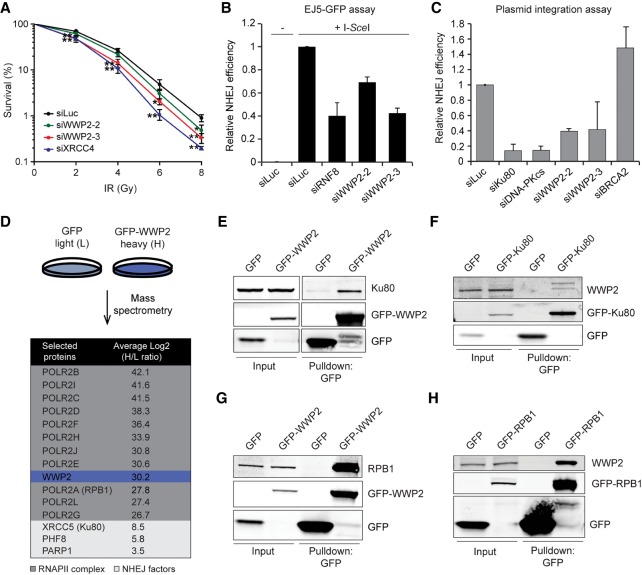
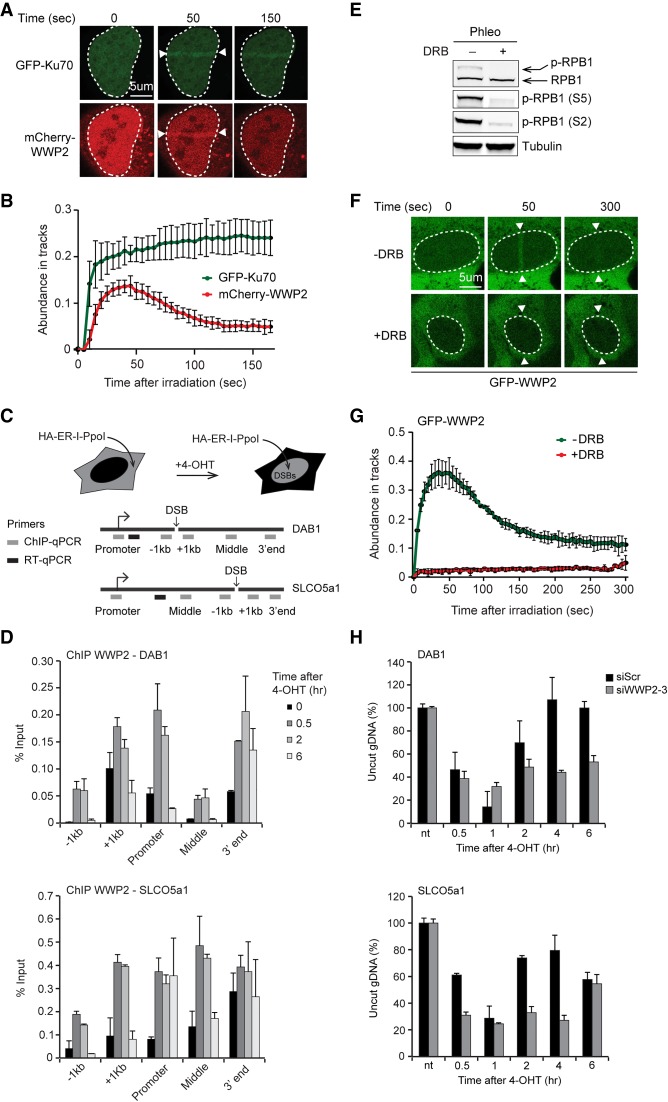
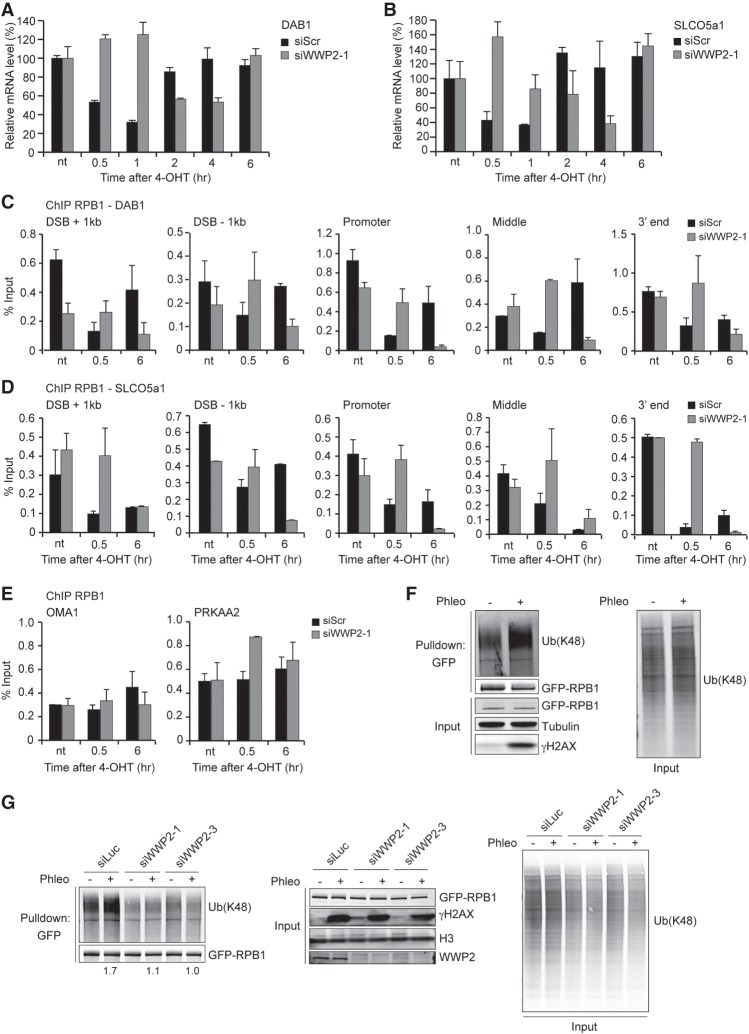
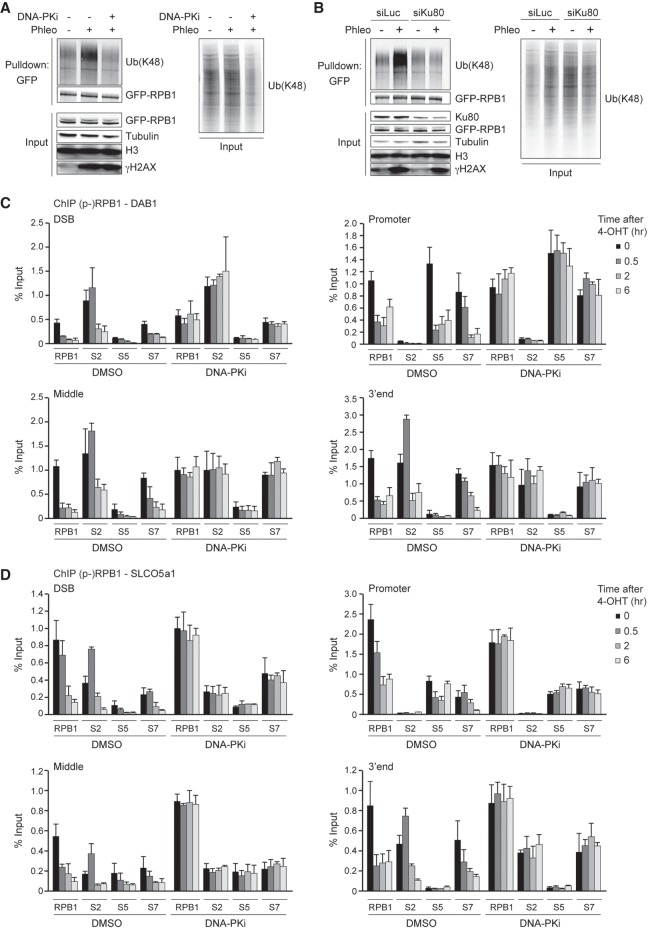
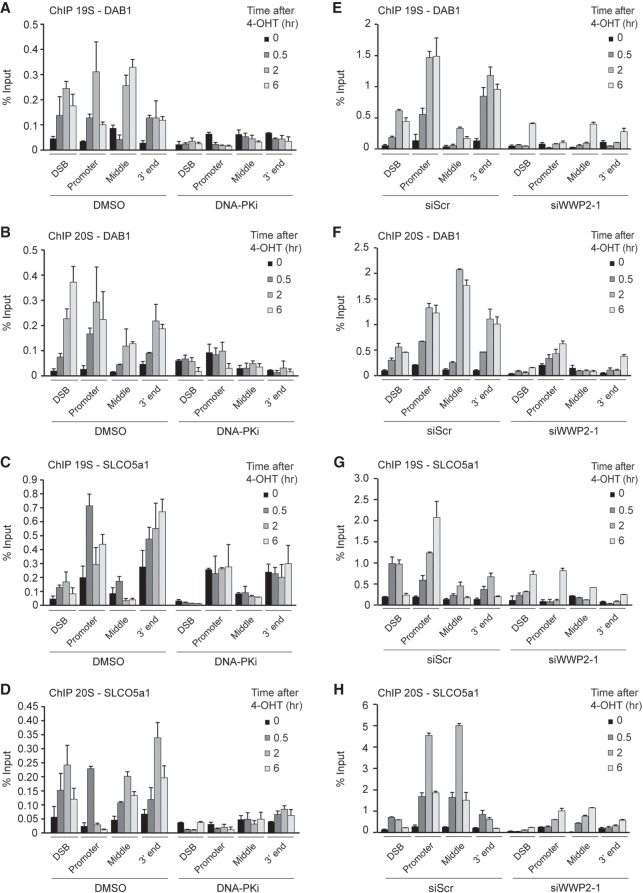
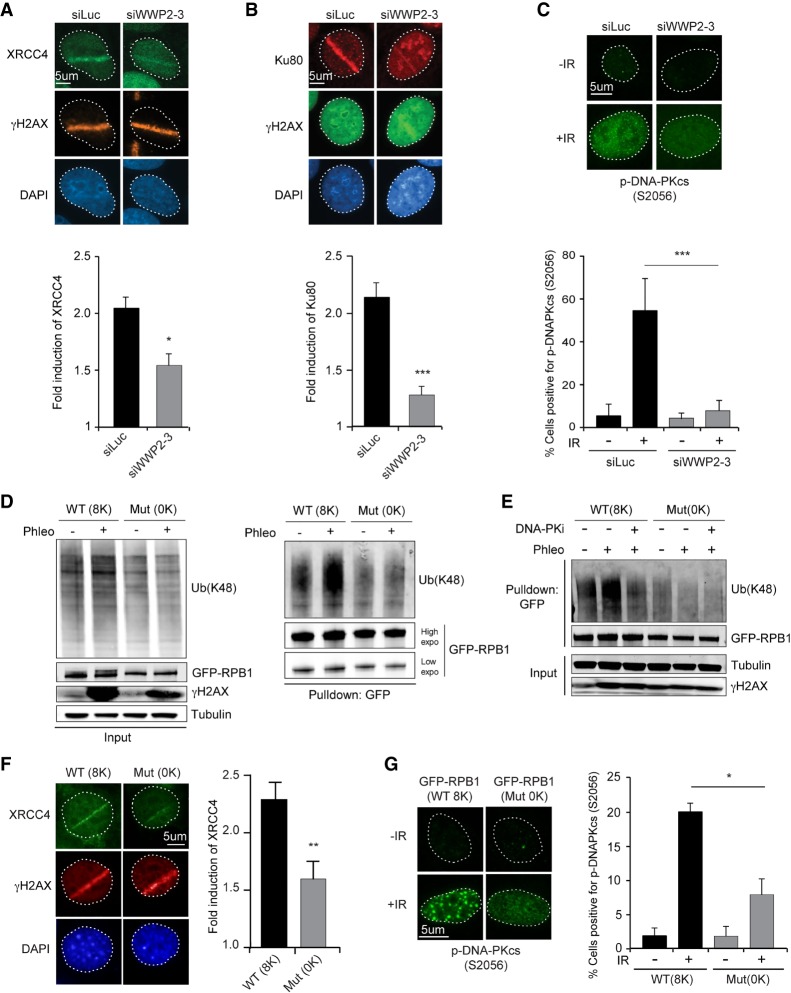
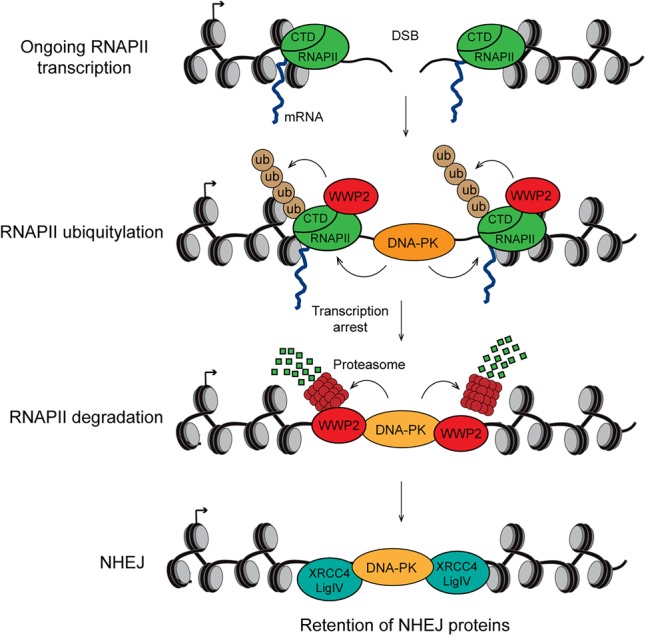
DNA double-strand breaks (DSBs) at RNA polymerase II (RNAPII) transcribed genes lead to inhibition of transcription. The DNA-dependent protein kinase (DNA-PK) complex plays a pivotal role in transcription inhibition at DSBs by stimulating proteasome-dependent eviction of RNAPII at these lesions. How DNA-PK triggers RNAPII eviction to inhibit transcription at DSBs remains unclear. Here we show that the HECT E3 ubiquitin ligase WWP2 associates with components of the DNA-PK and RNAPII complexes and is recruited to DSBs at RNAPII transcribed genes. In response to DSBs, WWP2 targets the RNAPII subunit RPB1 for K48-linked ubiquitylation, thereby driving DNA-PK- and proteasome-dependent eviction of RNAPII. The lack of WWP2 or expression of nonubiquitylatable RPB1 abrogates the binding of nonhomologous end joining (NHEJ) factors, including DNA-PK and XRCC4/DNA ligase IV, and impairs DSB repair. These findings suggest that WWP2 operates in a DNA-PK-dependent shutoff circuitry for RNAPII clearance that promotes DSB repair by protecting the NHEJ machinery from collision with the transcription machinery.
Keywords: DNA double-strand break repair; DNA-PK; RNAPII ubiquitylation; WWP2 HECT E3 ubiquitin ligase; transcription silencing.
© 2019 Caron et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials