

HCF-2 inhibits cell proliferation and activates differentiation-gene expression programs
- PMID: 31049581
- PMCID: PMC6582346
- DOI: 10.1093/nar/gkz307
HCF-2 inhibits cell proliferation and activates differentiation-gene expression programs
Abstract
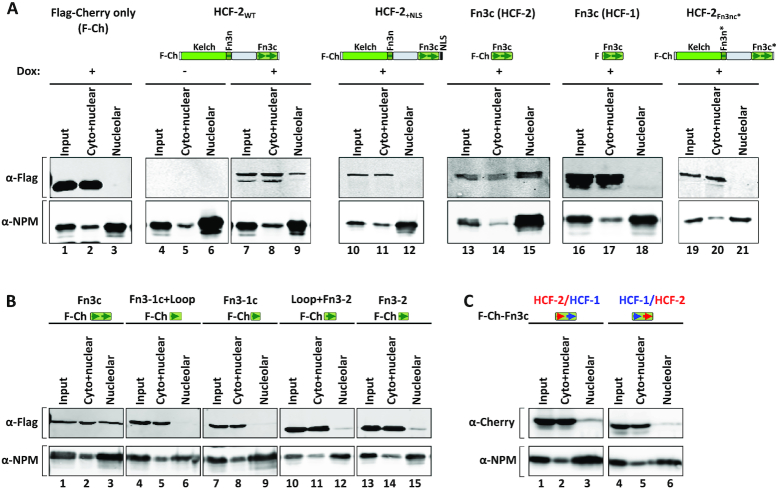
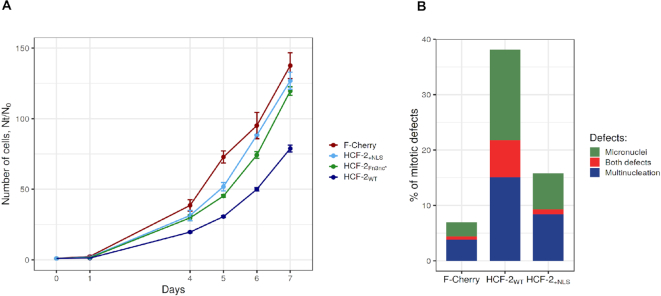
HCF-2 is a member of the host-cell-factor protein family, which arose in early vertebrate evolution as a result of gene duplication. Whereas its paralog, HCF-1, is known to act as a versatile chromatin-associated protein required for cell proliferation and differentiation, much less is known about HCF-2. Here, we show that HCF-2 is broadly present in human and mouse cells, and possesses activities distinct from HCF-1. Unlike HCF-1, which is excluded from nucleoli, HCF-2 is nucleolar-an activity conferred by one and a half C-terminal Fibronectin type 3 repeats and inhibited by the HCF-1 nuclear localization signal. Elevated HCF-2 synthesis in HEK-293 cells results in phenotypes reminiscent of HCF-1-depleted cells, including inhibition of cell proliferation and mitotic defects. Furthermore, increased HCF-2 levels in HEK-293 cells lead to inhibition of cell proliferation and metabolism gene-expression programs with parallel activation of differentiation and morphogenesis gene-expression programs. Thus, the HCF ancestor appears to have evolved into a small two-member protein family possessing contrasting nuclear versus nucleolar localization, and cell proliferation and differentiation functions.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures







References
-
- Zhang J. Evolution by gene duplication: an update. Trends Ecol. Evol. 2003; 18:292–298.
-
- Conant G.C., Wolfe K.H.. Turning a hobby into a job: how duplicated genes find new functions. Nat. Rev. Genet. 2008; 12:938–950. - PubMed
-
- Taylor J.S., Raes J.. Duplication and divergence: the evolution of new genes and old ideas. Annu. Rev. Genet. 2004; 38:615–643. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
