

Coxsackievirus B3 Responds to Polyamine Depletion via Enhancement of 2A and 3C Protease Activity
- PMID: 31052199
- PMCID: PMC6563312
- DOI: 10.3390/v11050403
Coxsackievirus B3 Responds to Polyamine Depletion via Enhancement of 2A and 3C Protease Activity
Abstract
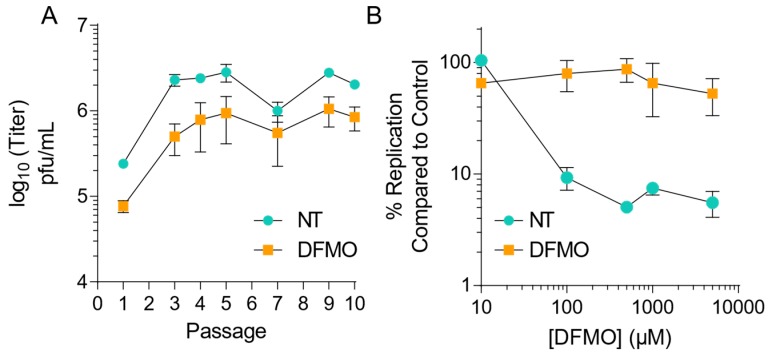
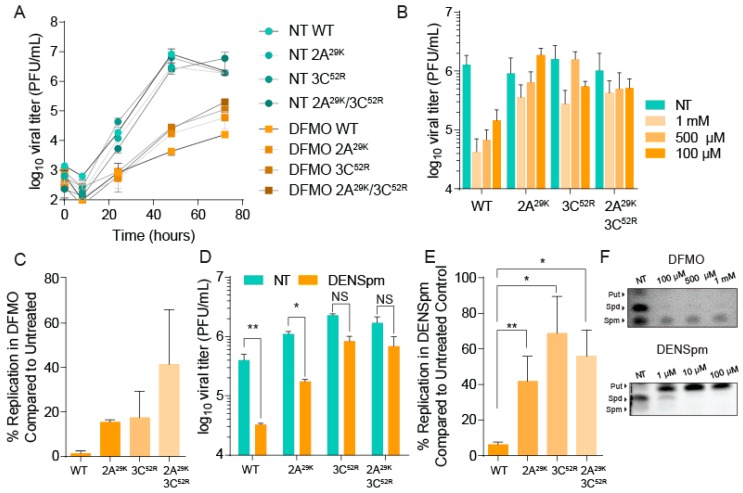
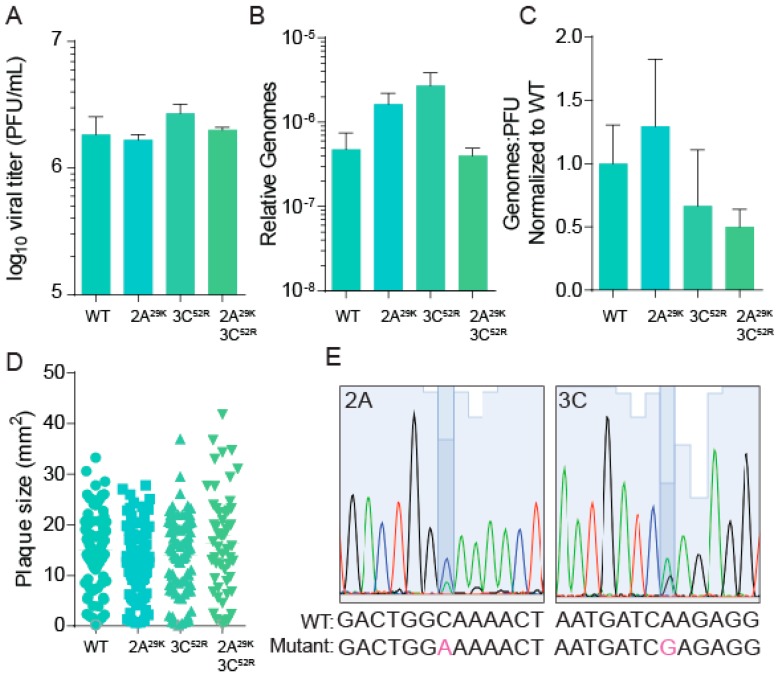
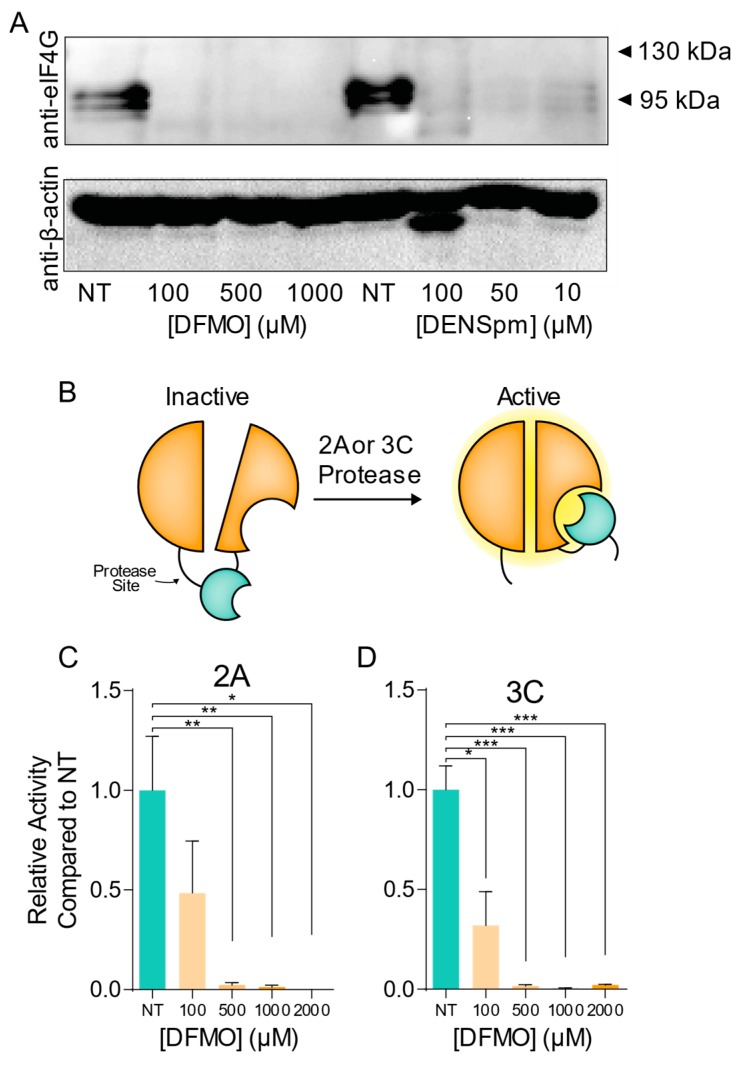
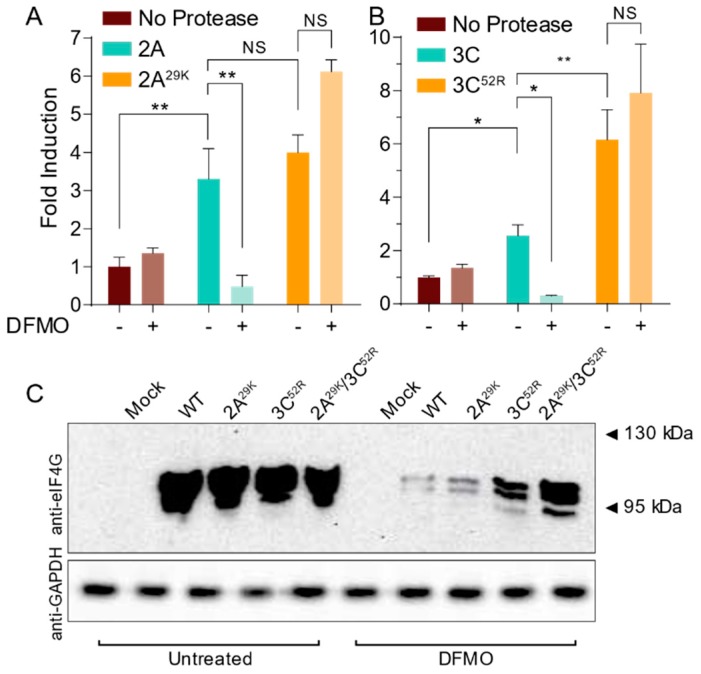
Polyamines are small positively-charged molecules abundant in eukaryotic cells that are crucial to RNA virus replication. In eukaryotic cells, polyamines facilitate processes such as transcription, translation, and DNA replication, and viruses similarly rely on polyamines to facilitate transcription and translation. Whether polyamines function at additional stages in viral replication remains poorly understood. Picornaviruses, including Coxsackievirus B3 (CVB3), are sensitive to polyamine depletion both in vitro and in vivo; however, precisely how polyamine function in picornavirus infection has not been described. Here, we describe CVB3 mutants that arise with passage in polyamine-depleted conditions. We observe mutations in the 2A and 3C proteases, and we find that these mutant proteases confer resistance to polyamine depletion. Using a split luciferase reporter system to measure protease activity, we determined that polyamines facilitate viral protease activity. We further observe that the 2A and 3C protease mutations enhance reporter protease activity in polyamine-depleted conditions. Finally, we find that these mutations promote cleavage of cellular eIF4G during infection of polyamine-depleted cells. In sum, our results suggest that polyamines are crucial to protease function during picornavirus infection. Further, these data highlight viral proteases as potential antiviral targets and highlight how CVB3 may overcome polyamine-depleting antiviral therapies.
Keywords: Coxsackievirus B3; polyamines; protease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Chapman N.M., Kim K.-S. Group B Coxsackieviruses. Springer; Berlin/Heidelberg, Germany: 2008. Persistent Coxsackievirus Infection: Enterovirus Persistence in Chronic Myocarditis and Dilated Cardiomyopathy; pp. 275–292. Current Topics in Microbiology and Immunology. - PubMed
-
- Lyoo H., Dorobantu C.M., van der Schaar H.M., van Kuppeveld F.J.M. Modulation of proteolytic polyprotein processing by coxsackievirus mutants resistant to inhibitors targeting phosphatidylinositol-4-kinase IIIβ or oxysterol binding protein. Antiviral Res. 2017;147:86–90. doi: 10.1016/j.antiviral.2017.10.006. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
