

Mitochondrial Flexibility of Breast Cancers: A Growth Advantage and a Therapeutic Opportunity
- PMID: 31052256
- PMCID: PMC6562467
- DOI: 10.3390/cells8050401
Mitochondrial Flexibility of Breast Cancers: A Growth Advantage and a Therapeutic Opportunity
Abstract
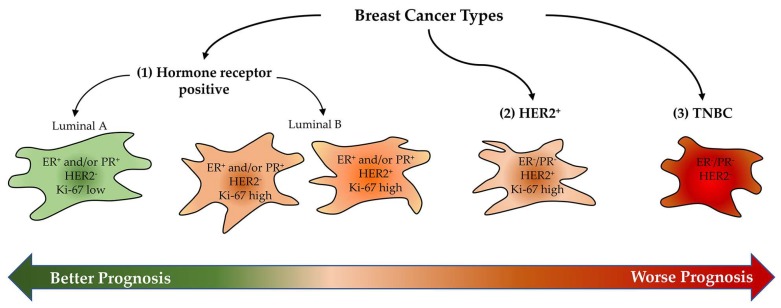
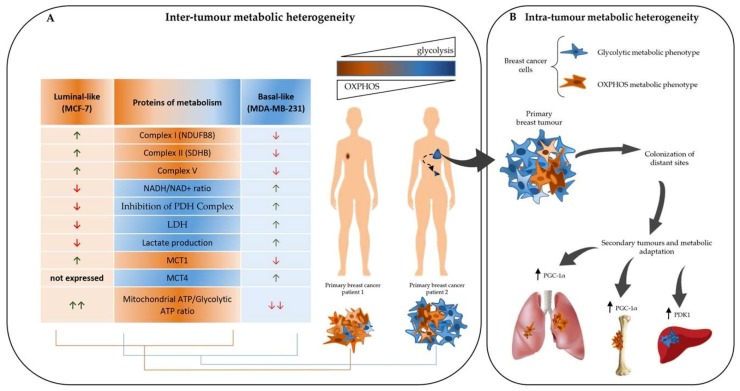
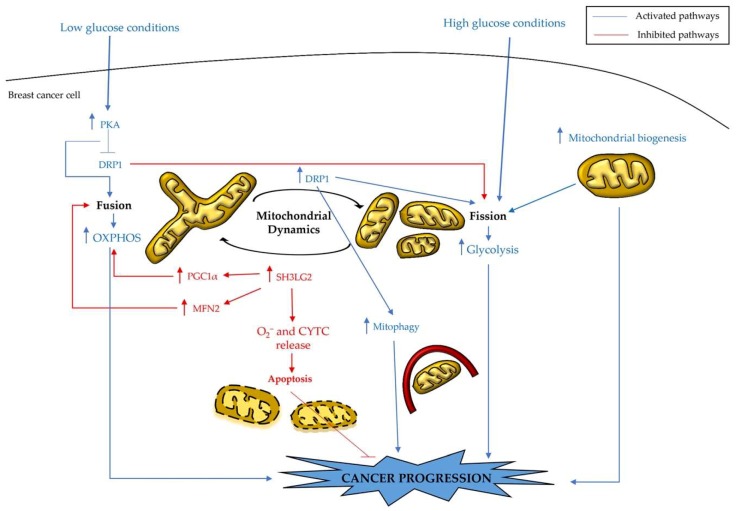
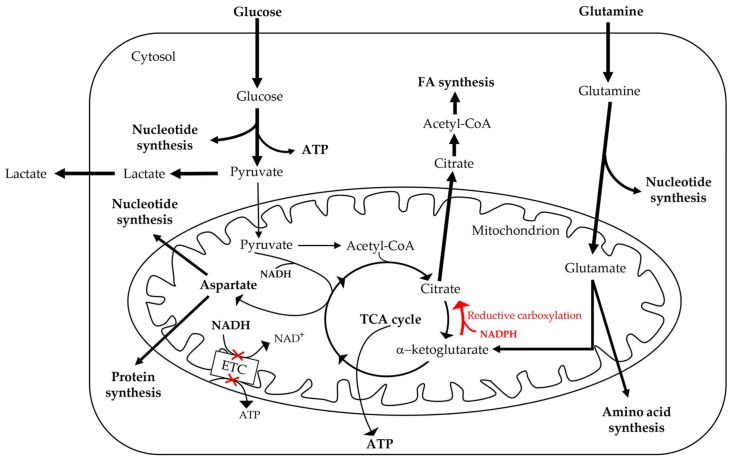
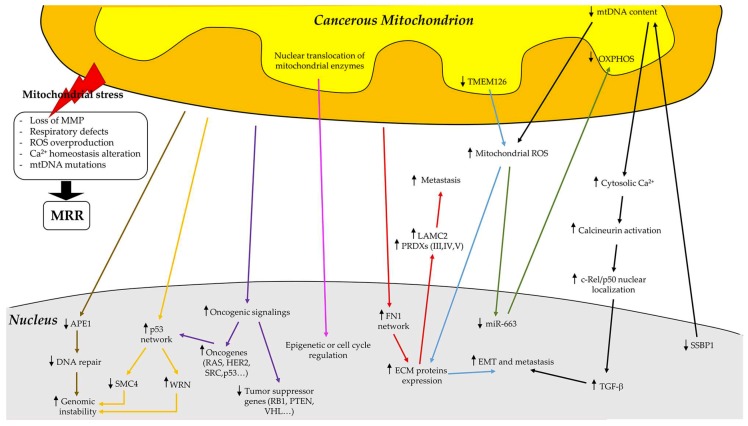
Breast cancers are very heterogeneous tissues with several cell types and metabolic pathways together sustaining the initiation and progression of disease and contributing to evasion from cancer therapies. Furthermore, breast cancer cells have an impressive metabolic plasticity that is regulated by the heterogeneous tumour microenvironment through bidirectional interactions. The structure and accessibility of nutrients within this unstable microenvironment influence the metabolism of cancer cells that shift between glycolysis and mitochondrial oxidative phosphorylation (OXPHOS) to produce adenosine triphosphate (ATP). In this scenario, the mitochondrial energetic pathways of cancer cells can be reprogrammed to modulate breast cancer's progression and aggressiveness. Moreover, mitochondrial alterations can lead to crosstalk between the mitochondria and the nucleus, and subsequently affect cancer tissue properties. This article reviewed the metabolic plasticity of breast cancer cells, focussing mainly on breast cancer mitochondrial metabolic reprogramming and the mitochondrial alterations influencing nuclear pathways. Finally, the therapeutic strategies targeting molecules and pathways regulating cancer mitochondrial alterations are highlighted.
Keywords: breast cancer; mitochondrial reprogramming; oxidative phosphorylation; therapeutic strategies; tumour microenvironment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ruocco M.R., Avagliano A., Granato G., Imparato V., Masone S., Masullo M., Nasso R., Montagnani S., Arcucci A. Involvement of Breast Cancer-Associated Fibroblasts in Tumor Development, Therapy Resistance and Evaluation of Potential Therapeutic Strategies. Curr. Med. Chem. 2018;25:3414–3434. doi: 10.2174/0929867325666180309120746. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
