

Distinct immunocyte-promoting and adipocyte-generating stromal components coordinate adipose tissue immune and metabolic tenors
- PMID: 31053654
- PMCID: PMC6648660
- DOI: 10.1126/sciimmunol.aaw3658
Distinct immunocyte-promoting and adipocyte-generating stromal components coordinate adipose tissue immune and metabolic tenors
Abstract
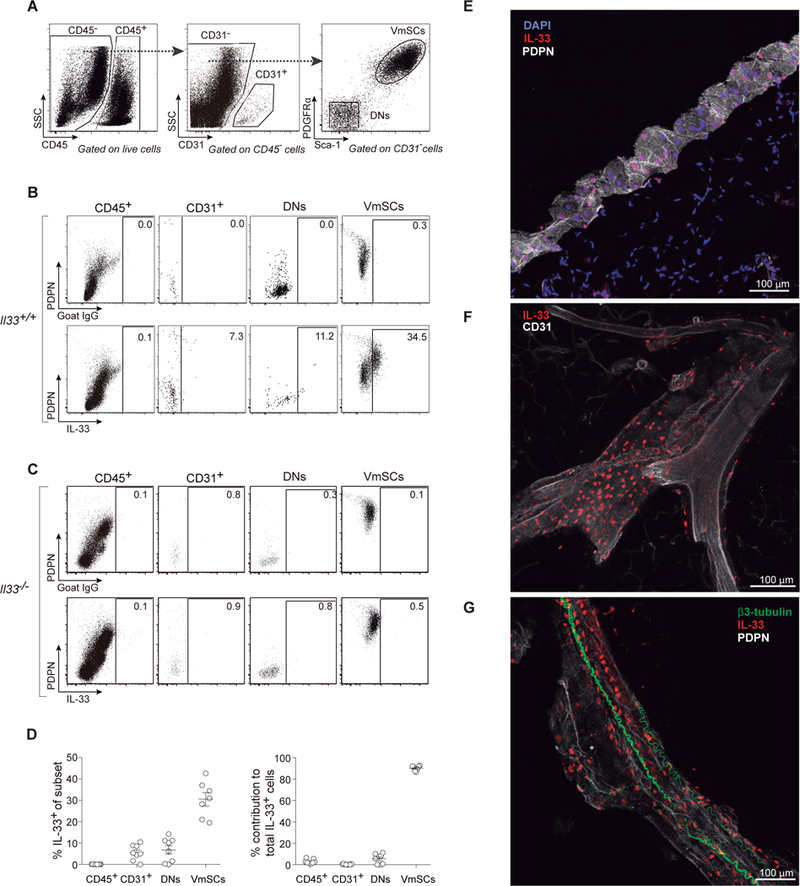
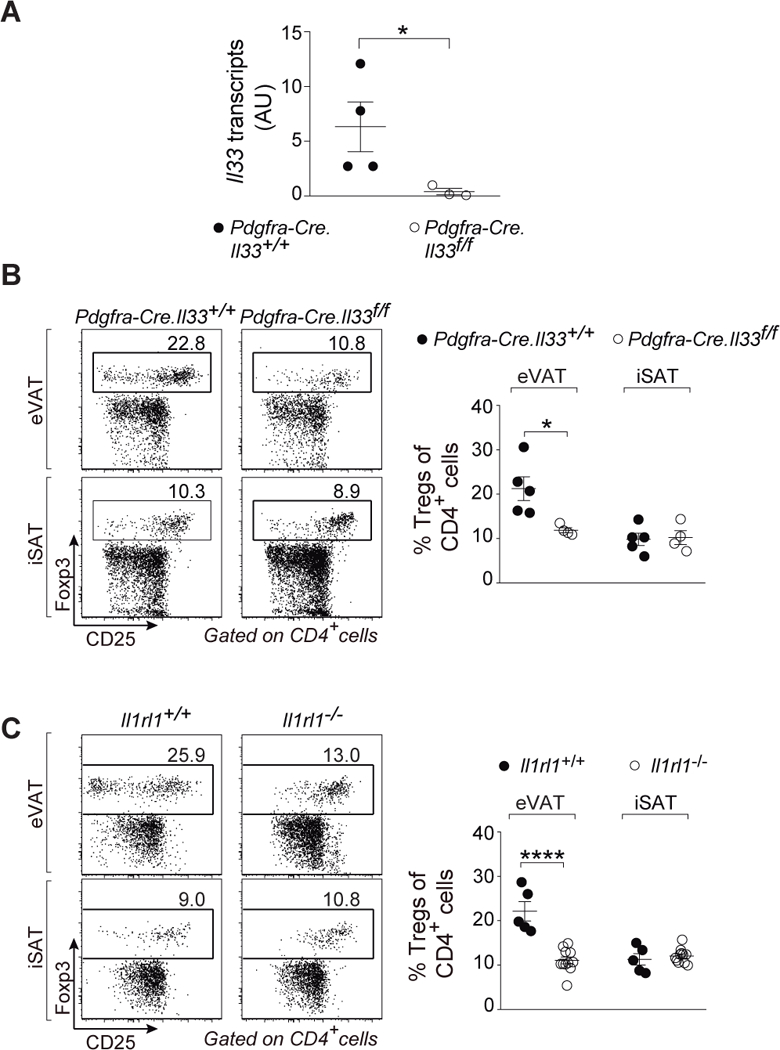
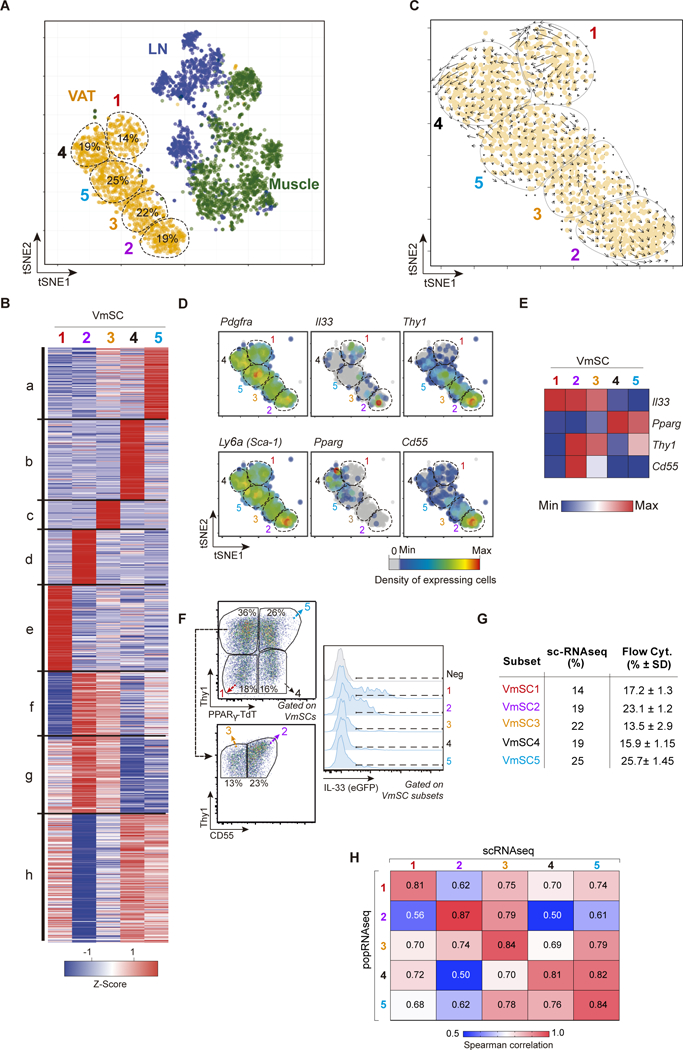
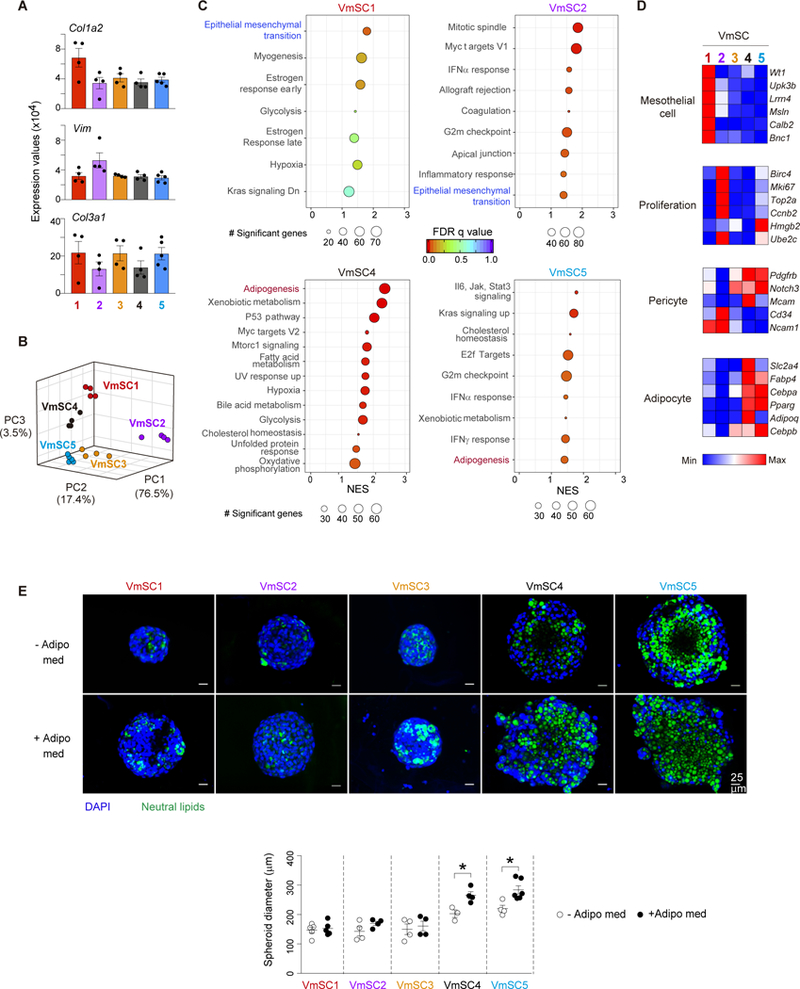
Regulatory T cells (Tregs) are key brakes on the visceral adipose tissue (VAT) inflammation that regulates local and systemic metabolic tenor. Breakdown of this regulation promotes type 2 diabetes. The cytokine IL-33 expands and sustains the unique Treg population residing within VAT. Here, relying on single-cell RNA sequencing, we identified the major IL-33 producers in VAT to be particular mesenchymal stromal cell subtypes, related to but distinct from adipocyte progenitor cells. We explored modulation of the VAT stromal cell landscape with physiologic variables such as age and sex, as well as its remodeling in pathogenic states like obesity. Last, we uncovered a VAT Treg:stromal cell negative regulatory loop that keeps the potent effect of IL-33 under rein.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
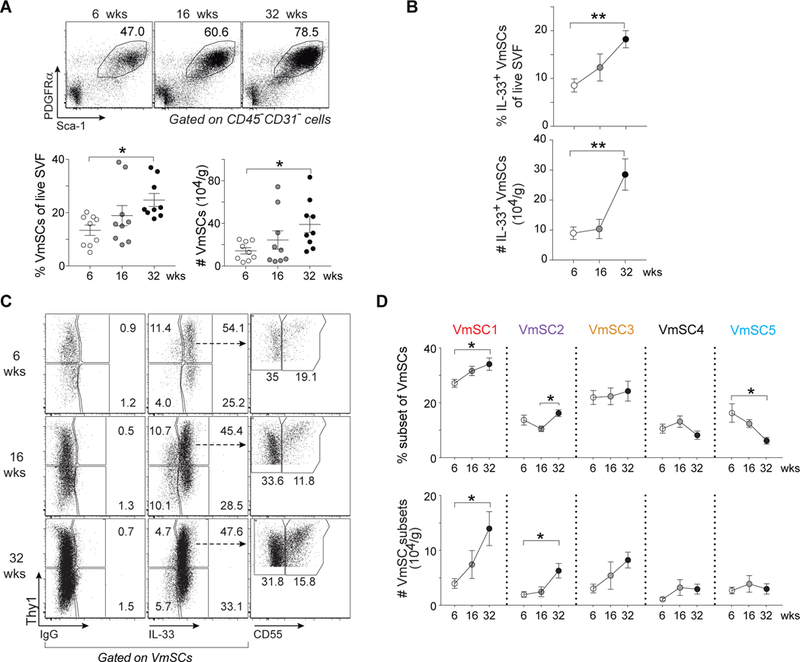
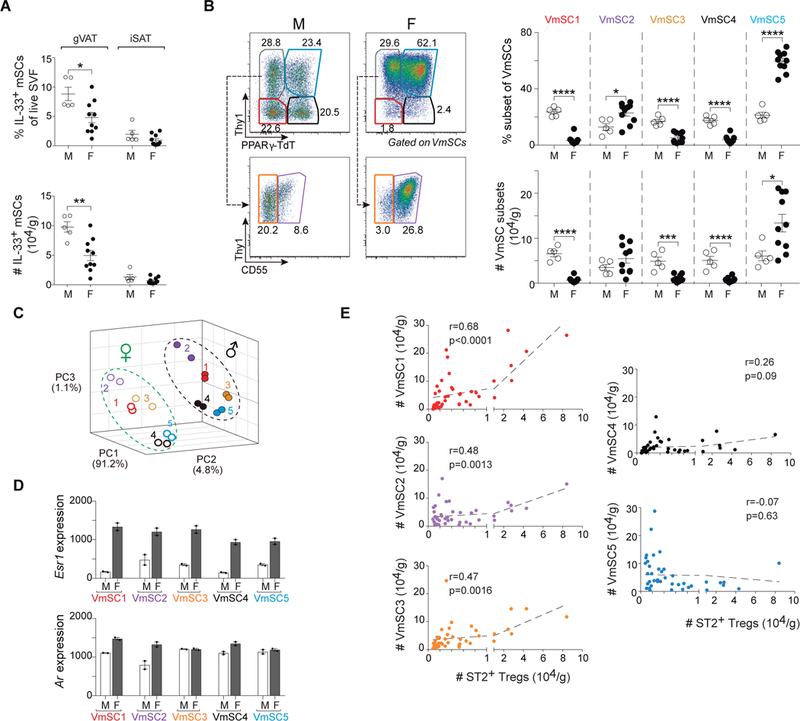
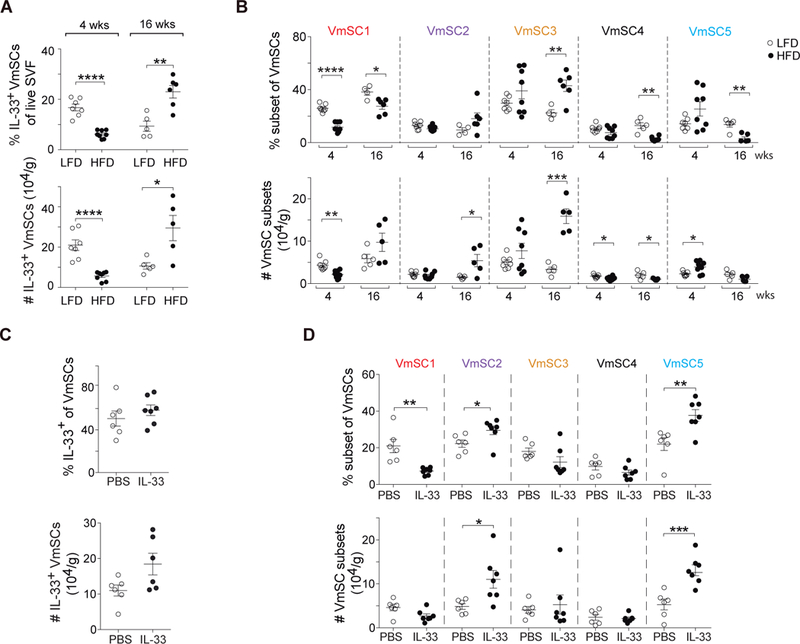
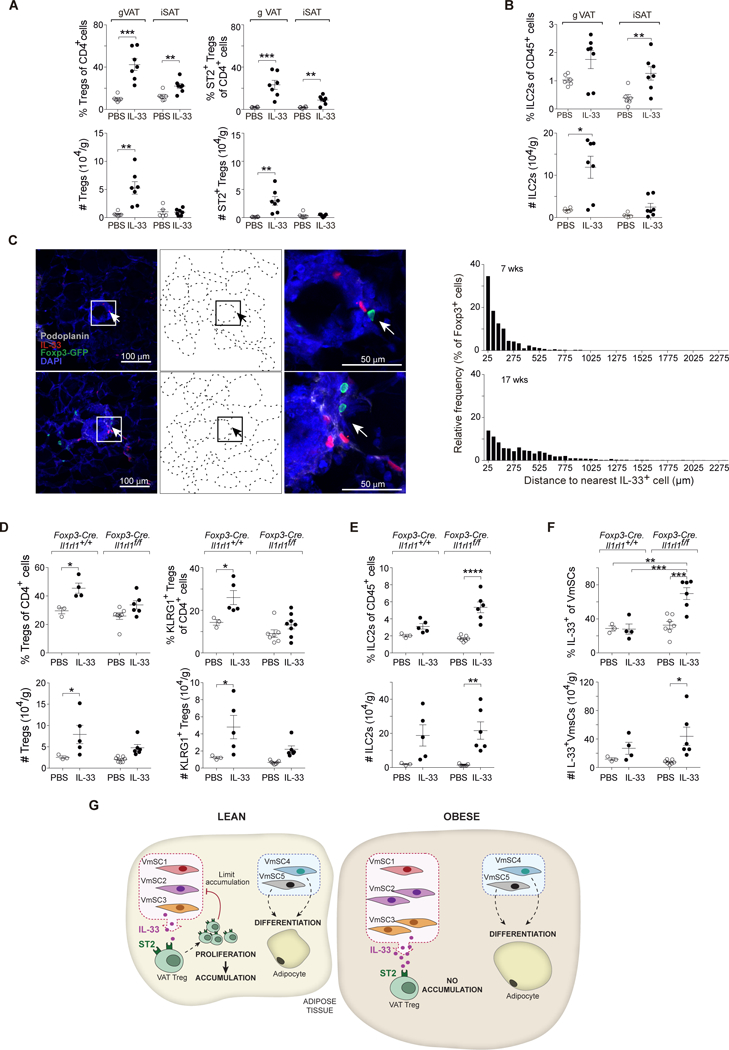
Comment in
-
Stromal IL-33 balances fat stores.Nat Rev Immunol. 2019 Jul;19(7):412-413. doi: 10.1038/s41577-019-0179-7. Nat Rev Immunol. 2019. PMID: 31110269 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
