

Crosstalk between RNA Pol II C-Terminal Domain Acetylation and Phosphorylation via RPRD Proteins
- PMID: 31054975
- PMCID: PMC6588463
- DOI: 10.1016/j.molcel.2019.04.008
Crosstalk between RNA Pol II C-Terminal Domain Acetylation and Phosphorylation via RPRD Proteins
Abstract
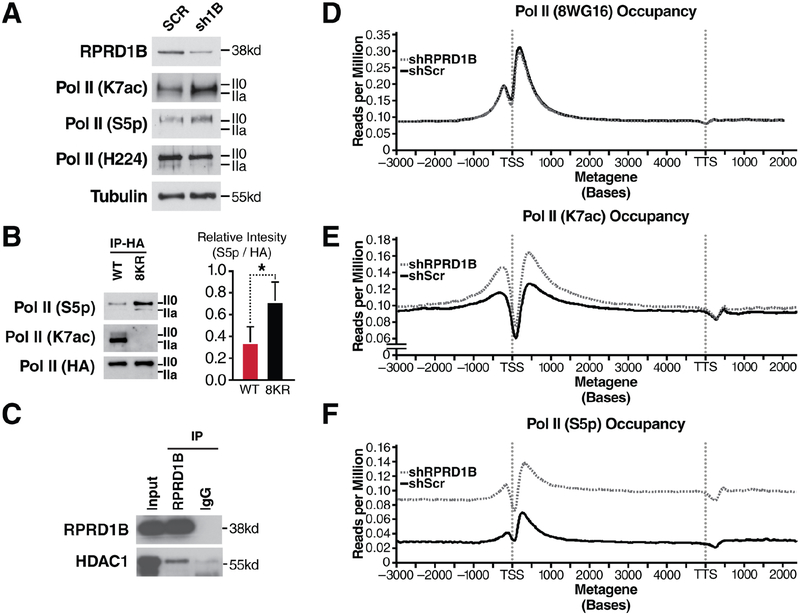
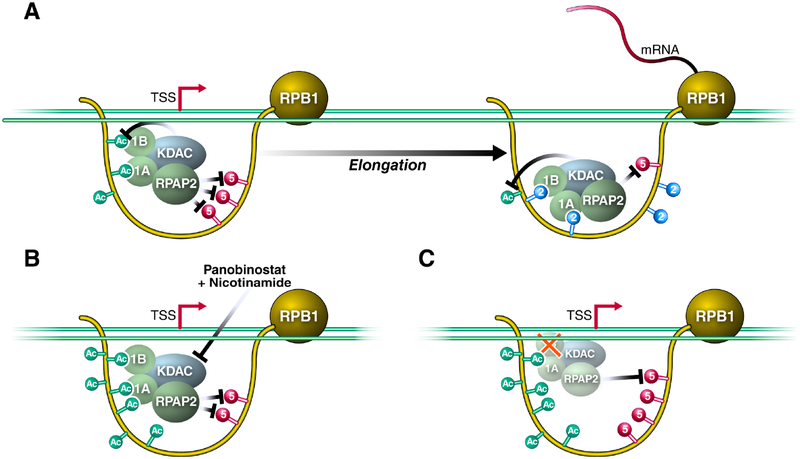
Post-translational modifications of the RNA polymerase II C-terminal domain (CTD) coordinate the transcription cycle. Crosstalk between different modifications is poorly understood. Here, we show how acetylation of lysine residues at position 7 of characteristic heptad repeats (K7ac)-only found in higher eukaryotes-regulates phosphorylation of serines at position 5 (S5p), a conserved mark of polymerases initiating transcription. We identified the regulator of pre-mRNA-domain-containing (RPRD) proteins as reader proteins of K7ac. K7ac enhanced CTD peptide binding to the CTD-interacting domain (CID) of RPRD1A and RPRD1B proteins in isothermal calorimetry and molecular modeling experiments. Deacetylase inhibitors increased K7ac- and decreased S5-phosphorylated polymerases, consistent with acetylation-dependent S5 dephosphorylation by an RPRD-associated S5 phosphatase. Consistent with this model, RPRD1B knockdown increased S5p but enhanced K7ac, indicating that RPRD proteins recruit K7 deacetylases, including HDAC1. We also report autoregulatory crosstalk between K7ac and S5p via RPRD proteins and their interactions with acetyl- and phospho-eraser proteins.
Keywords: C-terminal domain; Pol II CTD; RNA polymerase II; acetylation; crosstalk; gene regulation; histone deacetylase; phosphorylation; post-translational modification; transcription.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
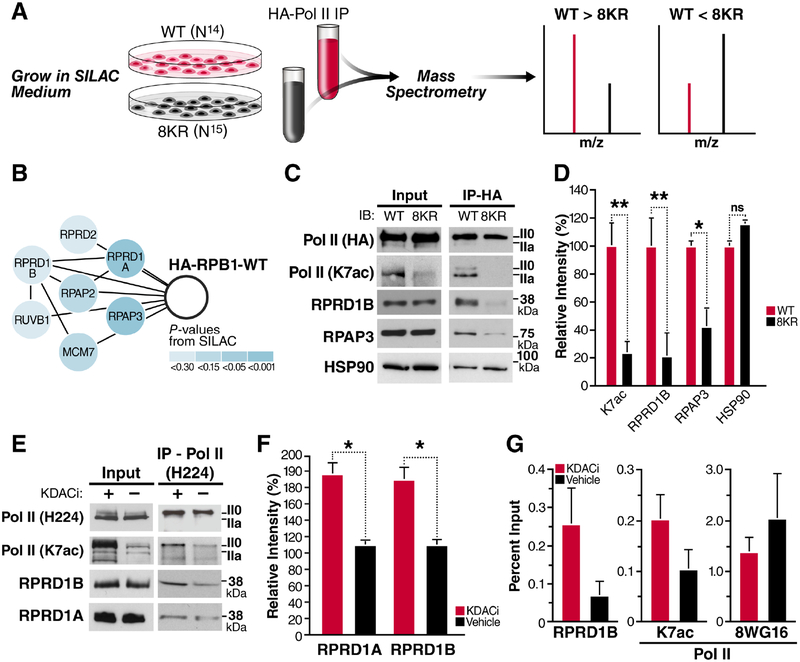
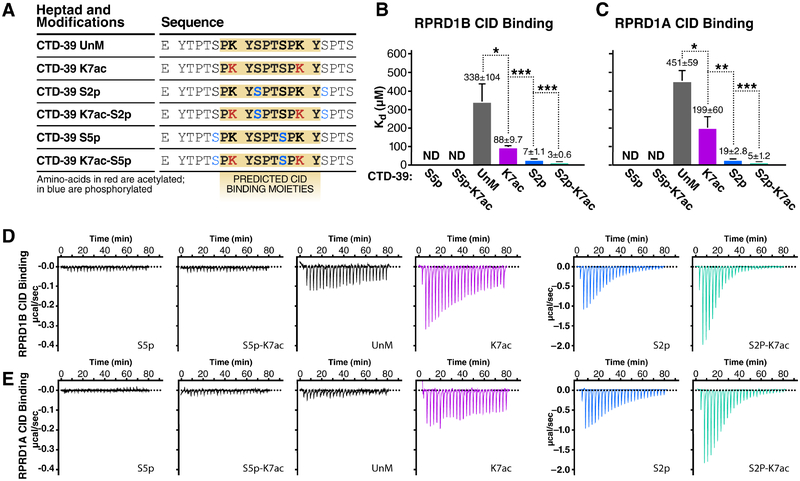
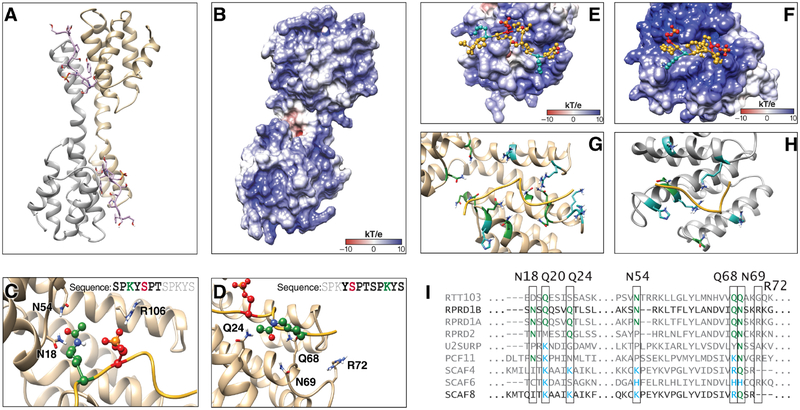
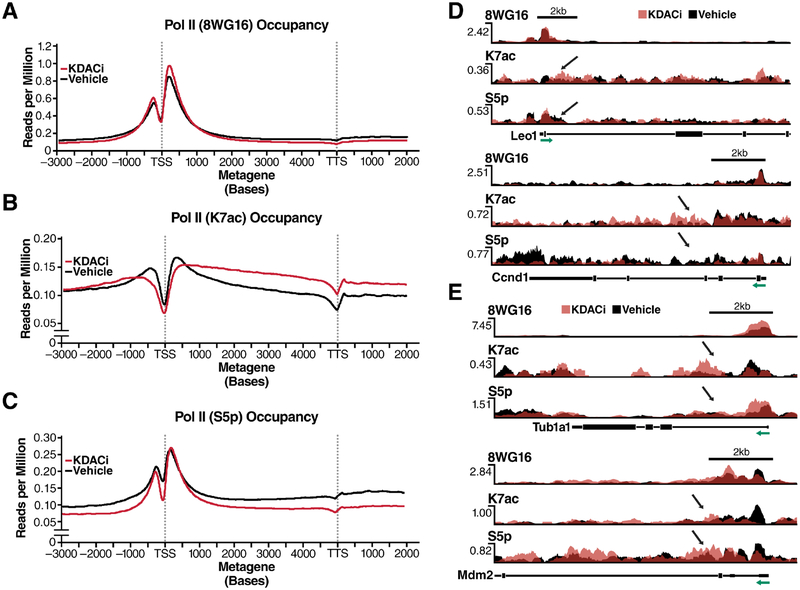
References
-
- BECKER R, LOLL B & MEINHART A 2008. Snapshots of the RNA processing factor SCAF8 bound to different phosphorylated forms of the carboxyl-terminal domain of RNA polymerase II. J Biol Chem, 283, 22659–69. - PubMed
-
- BROOKES E, DE SANTIAGO I, HEBENSTREIT D, MORRIS KJ, CARROLL T, XIE SQ, STOCK JK, HEIDEMANN M, EICK D, NOZAKI N, KIMURA H, RAGOUSSIS J, TEICHMANN SA & POMBO A 2012. Polycomb associates genome-wide with a specific RNA polymerase II variant, and regulates metabolic genes in ESCs. Cell Stem Cell, 10, 157–70. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
