

Allosteric Regulation of Oligomerization by a B12 Trafficking G-Protein Is Corrupted in Methylmalonic Aciduria
- PMID: 31056463
- PMCID: PMC6642015
- DOI: 10.1016/j.chembiol.2019.03.014
Allosteric Regulation of Oligomerization by a B12 Trafficking G-Protein Is Corrupted in Methylmalonic Aciduria
Abstract
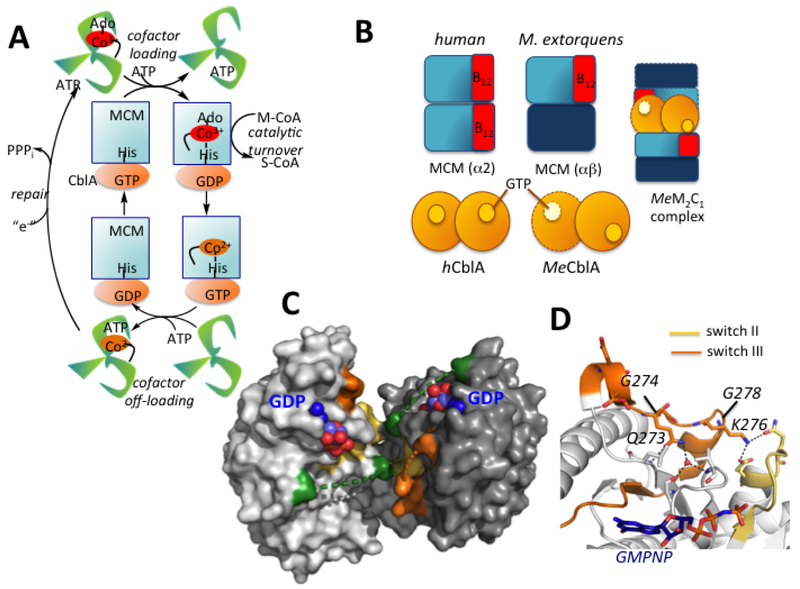
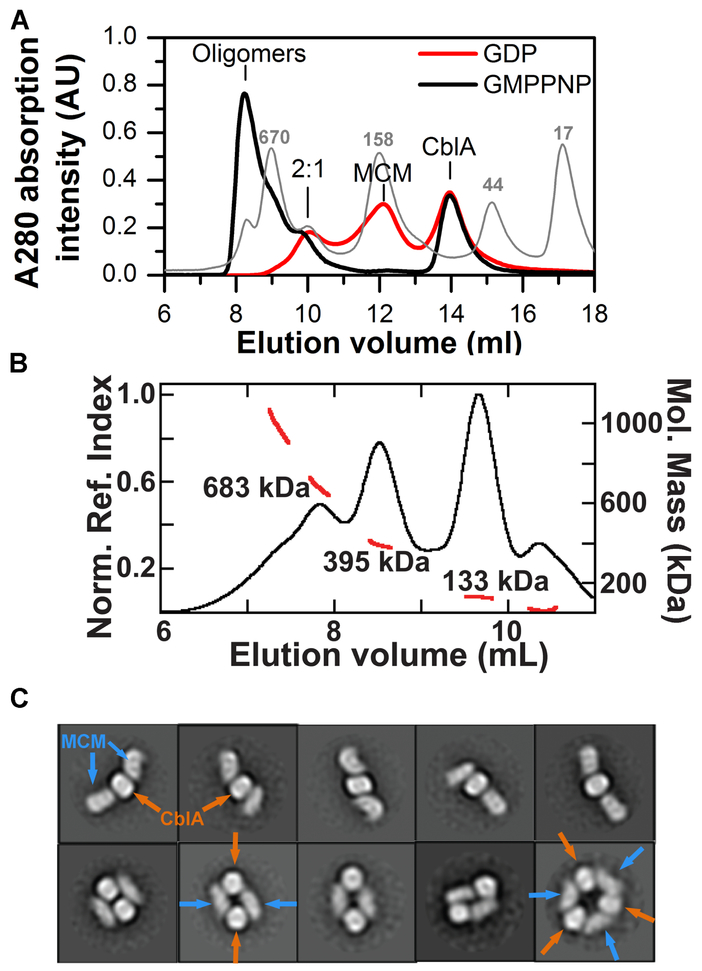
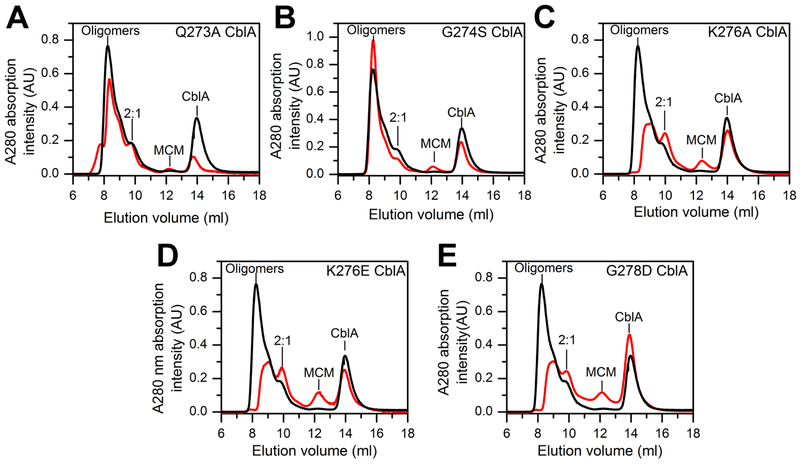
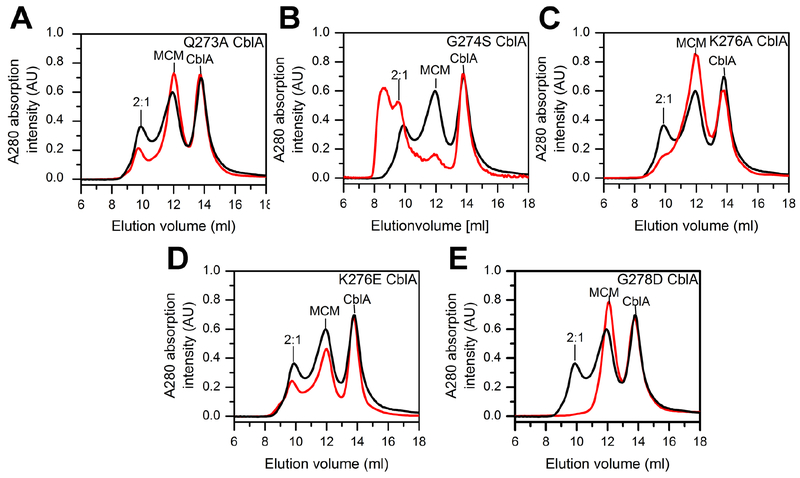
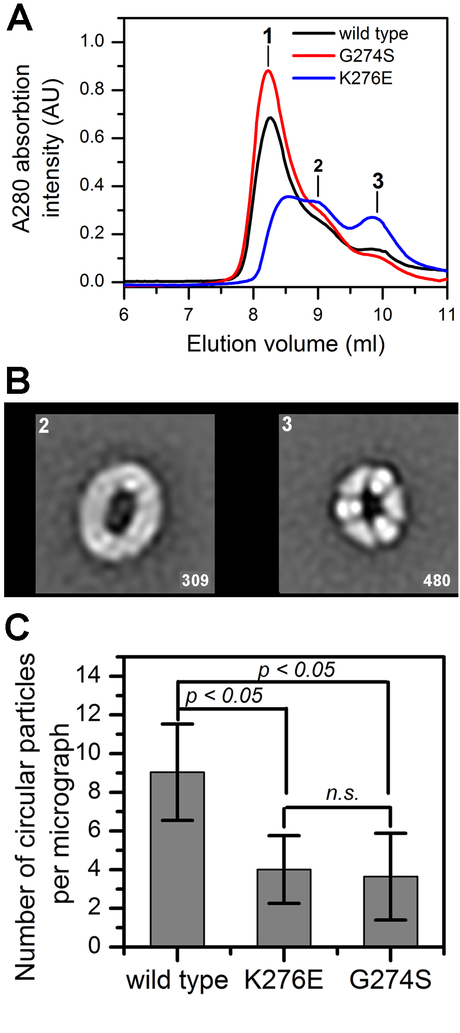
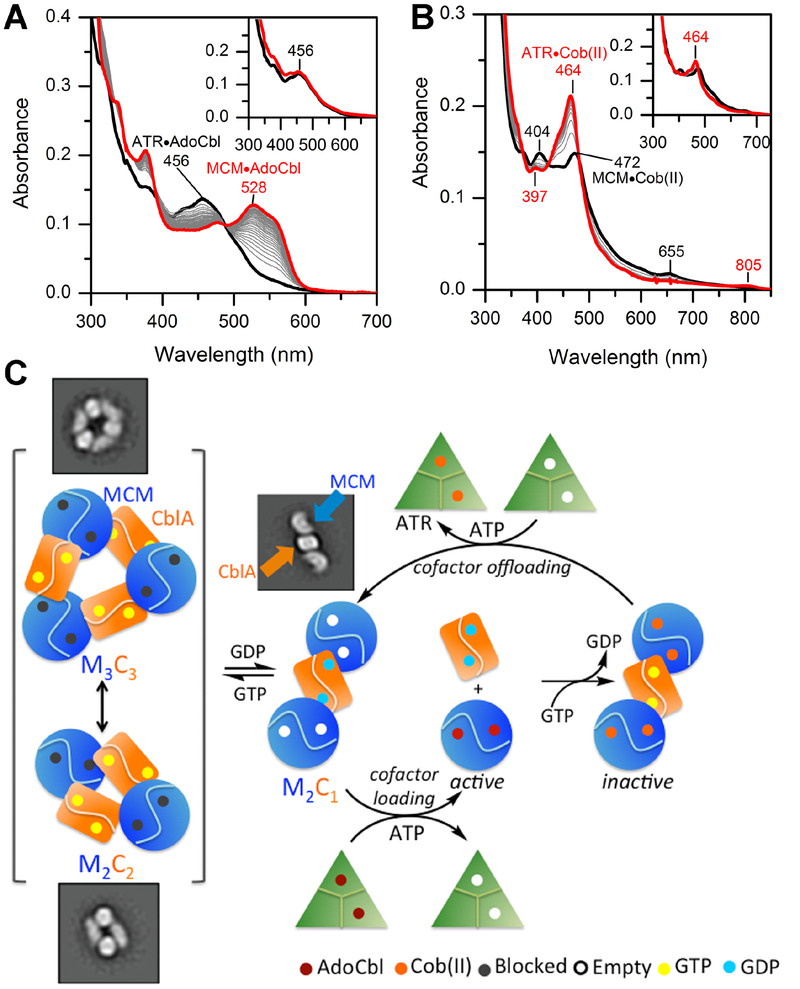
Allosteric regulation of methylmalonyl-CoA mutase (MCM) by the G-protein chaperone CblA is transduced via three "switch" elements that gate the movement of the B12 cofactor to and from MCM. Mutations in CblA and MCM cause hereditary methylmalonic aciduria. Unlike the bacterial orthologs used previously to model disease-causing mutations, human MCM and CblA exhibit a complex pattern of regulation that involves interconverting oligomers, which are differentially sensitive to the presence of GTP versus GDP. Patient mutations in the switch III region of CblA perturb the nucleotide-sensitive distribution of the oligomeric complexes with MCM, leading to loss of regulated movement of B12 to and/or from MCM and explain the molecular mechanism of the resulting disease.
Keywords: G-protein; GTPase; MMAA; cblA; cobalamin; cofactor; metal trafficking; metalloprotein; trafficking; vitamin B(12).
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interest
The authors declare that no competing financial interests exist.
Figures
References
-
- Banerjee R (2003). Radical carbon skeleton rearrangements: catalysis by coenzyme B12-dependent mutases. Chem Rev 103, 2083–2094. - PubMed
-
- Breinig S, Kervinen J, Stith L, Wasson AS, Fairman R, Wlodawer A, Zdanov A, and Jaffe EK (2003). Control of tetrapyrrole biosynthesis by alternate quaternary forms of porphobilinogen synthase. Nat Struct Biol 10, 757–763. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
