

Non-canonical Notch Signaling Regulates Actin Remodeling in Cell Migration by Activating PI3K/AKT/Cdc42 Pathway
- PMID: 31057403
- PMCID: PMC6477508
- DOI: 10.3389/fphar.2019.00370
Non-canonical Notch Signaling Regulates Actin Remodeling in Cell Migration by Activating PI3K/AKT/Cdc42 Pathway
Abstract
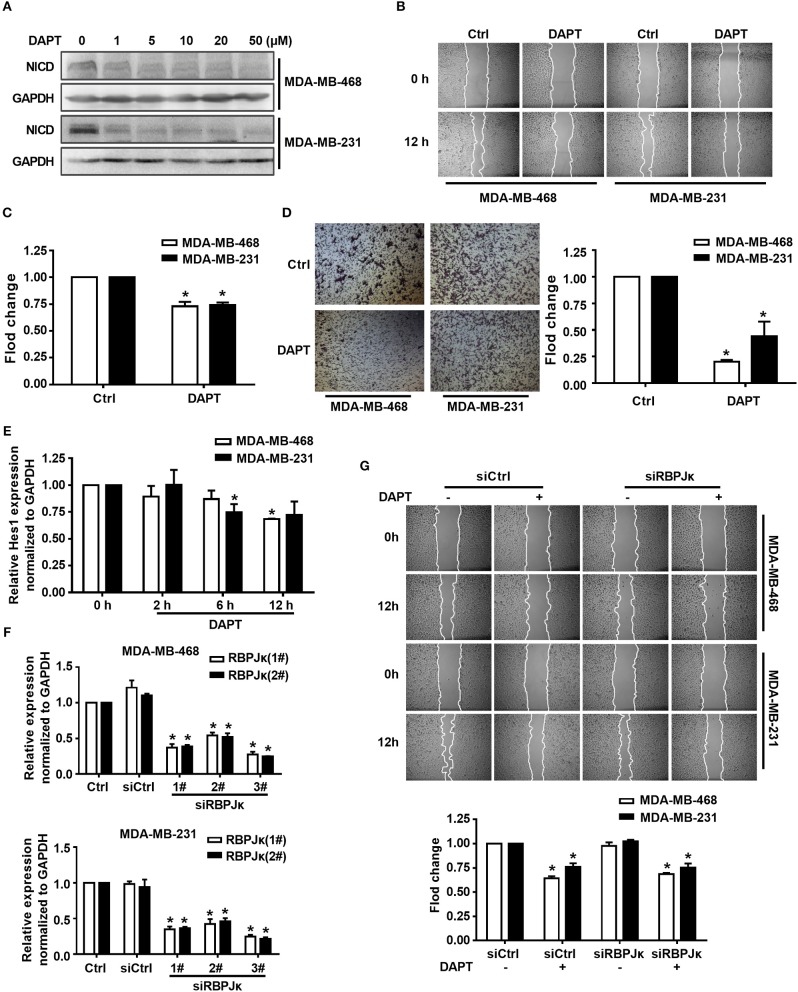
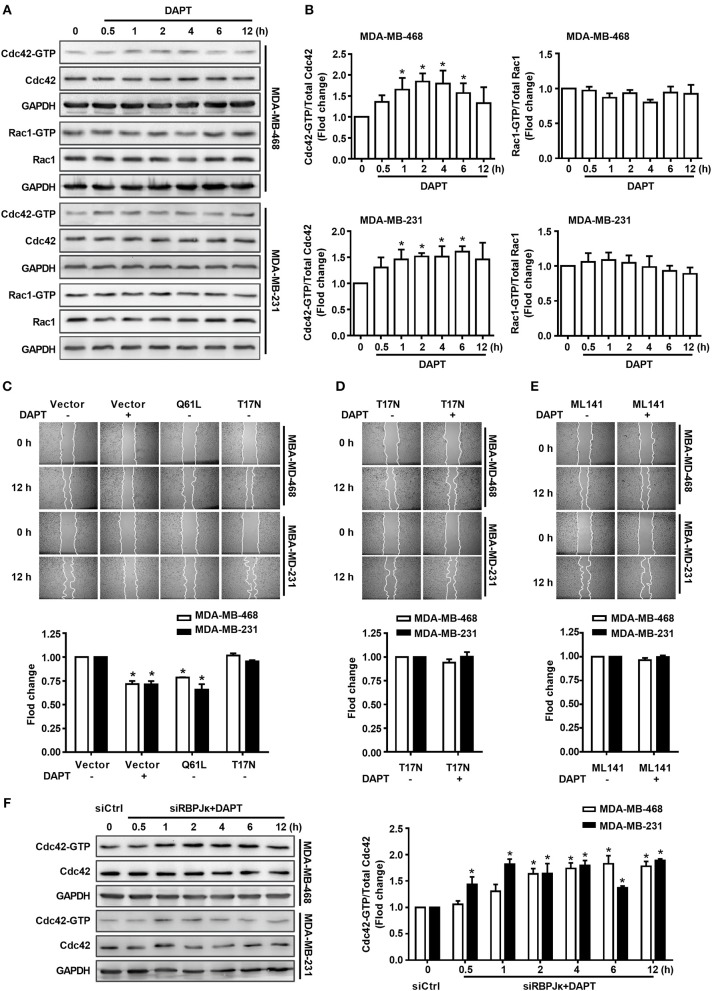
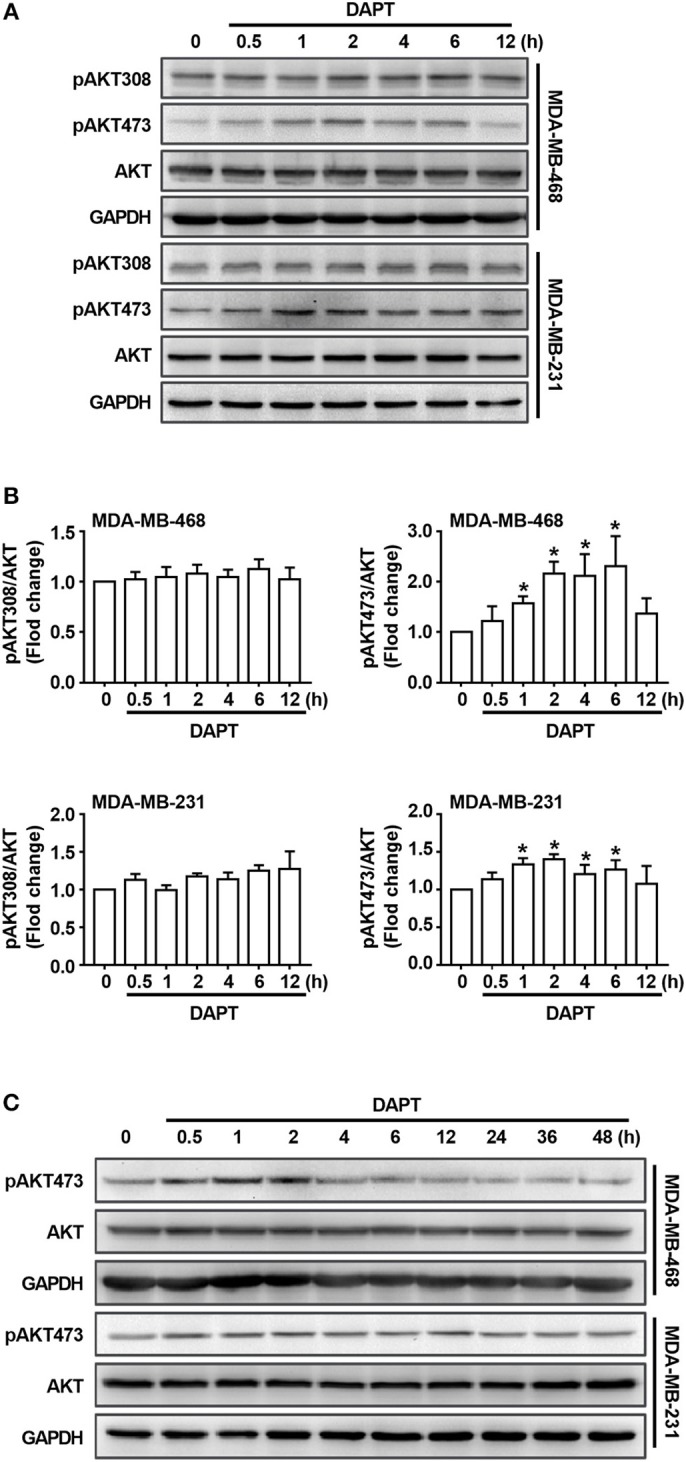
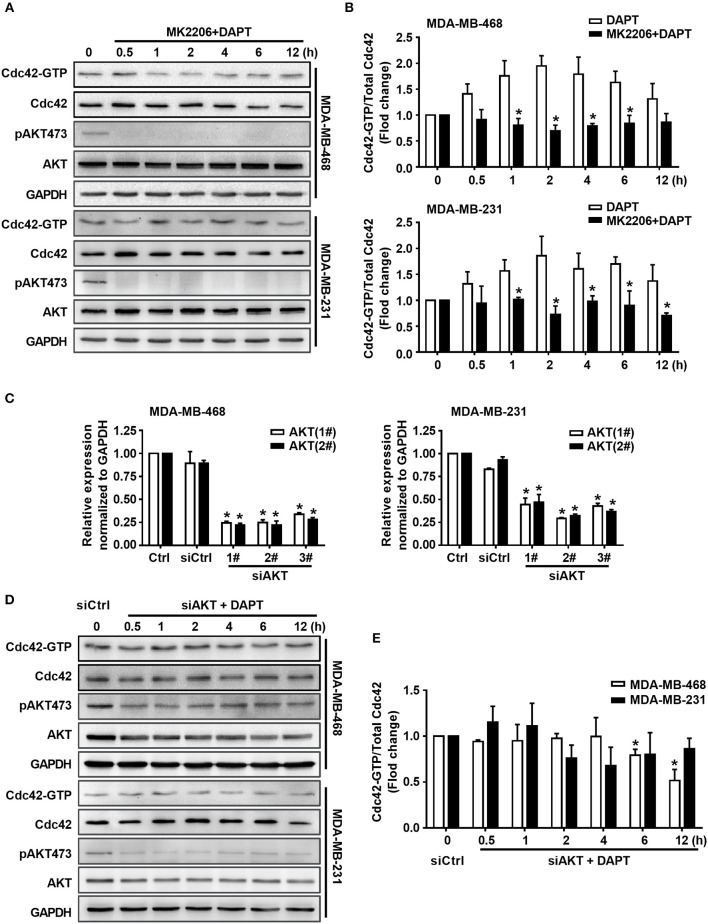
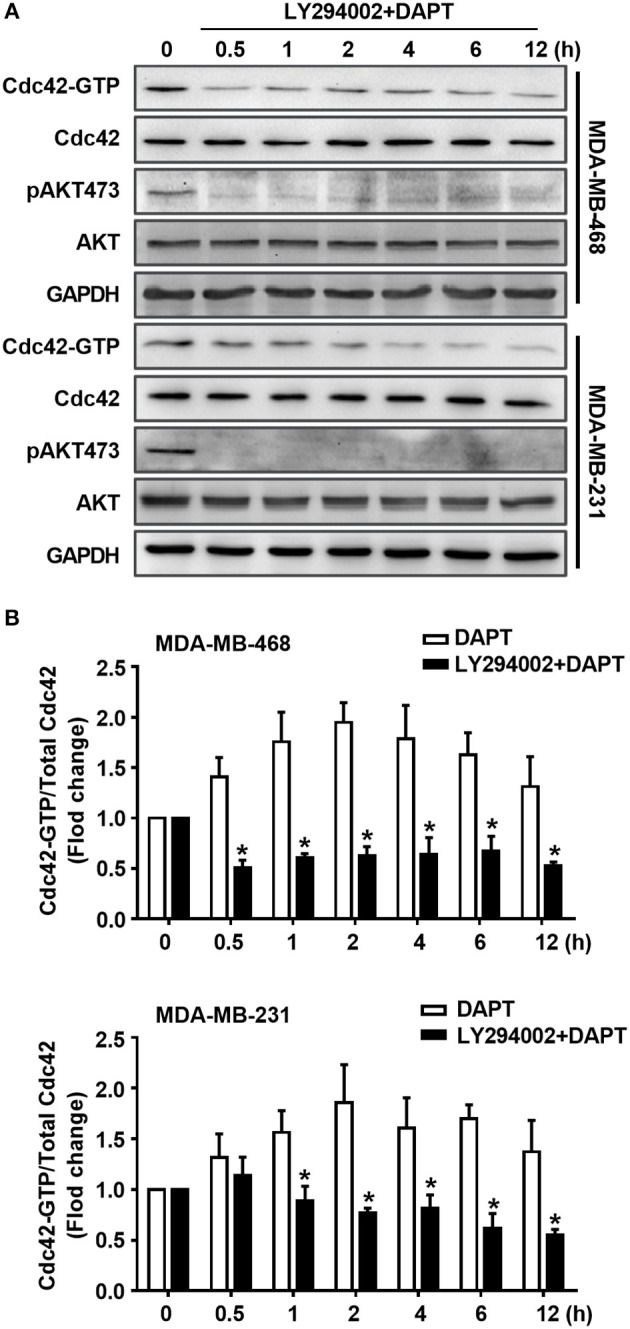
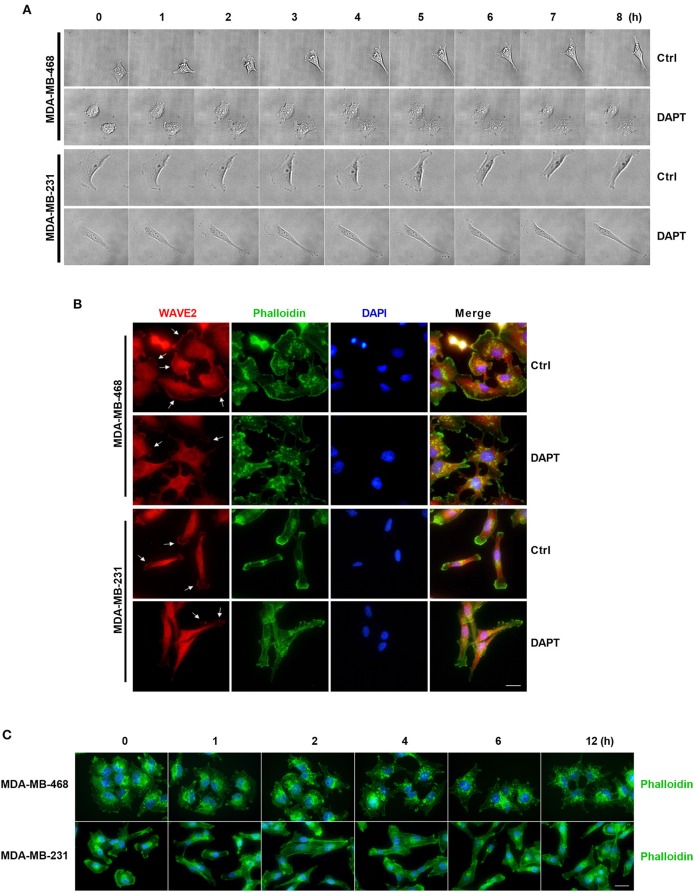
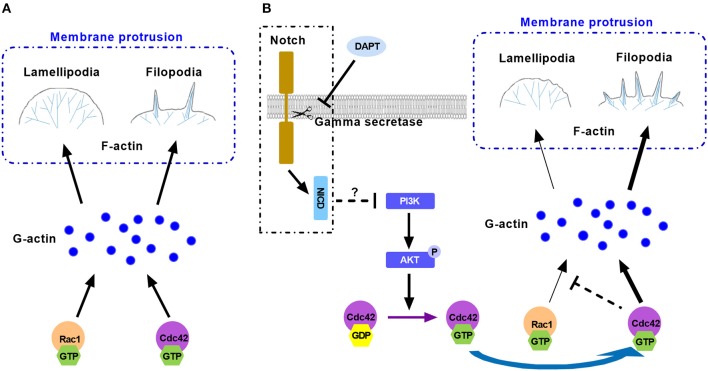
Tumor cell migration is a critical step in cancer metastasis. Over-activated Notch pathway can promote the migration of cancer cells, especially in the breast cancer. However, the underlying mechanism of non-canonical Notch signaling in modulating the migration has not yet been clearly characterized. Here we demonstrated that DAPT, a gamma secretase inhibitor, inhibited protrusion formation and cell motility, and then reduced the migration of triple-negative breast cancer cells, through increasing the activity of Cdc42 by non-canonical Notch pathway. Phosphorylation of AKT on S473 was surprisingly increased when Notch signaling was inhibited by DAPT. Inhibition of PI3K and AKT by LY294002 and MK2206, respectively, or knockdown of AKT expression by siRNA blocked DAPT-induced activation of Cdc42. Moreover, immunofluorescence staining further showed that DAPT treatment reduced the formation of lamellipodia and induced actin cytoskeleton remodeling. Taken together, these results indicated that DAPT inhibited Notch signaling and consequently activated PI3K/AKT/Cdc42 signaling by non-canonical pathway, facilitated the formation of filopodia and inhibited the assembly of lamellipodia, and finally resulted in the decrease of migration activity of breast cancer cells.
Keywords: Cdc42; PI3K/AKT; actin cytoskeleton; membrane protrusions; non-canonical Notch.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous