

Citrobacter rodentium Induces Tissue-Resident Memory CD4+ T Cells
- PMID: 31061145
- PMCID: PMC6589064
- DOI: 10.1128/IAI.00295-19
Citrobacter rodentium Induces Tissue-Resident Memory CD4+ T Cells
Abstract
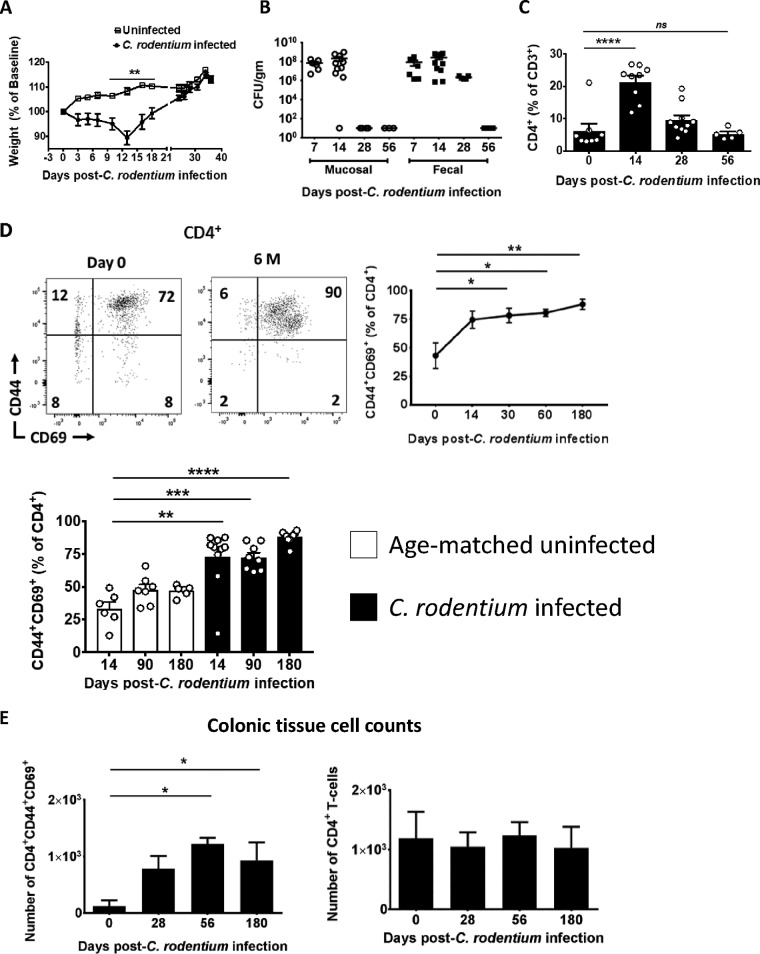
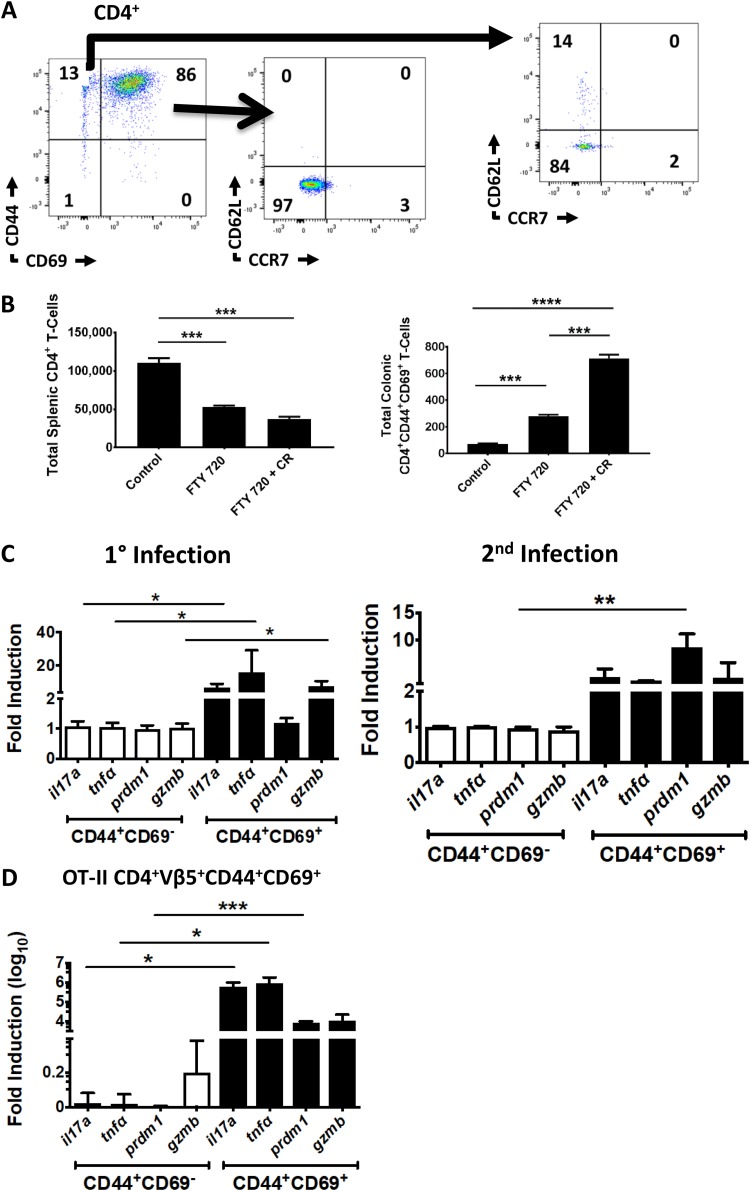
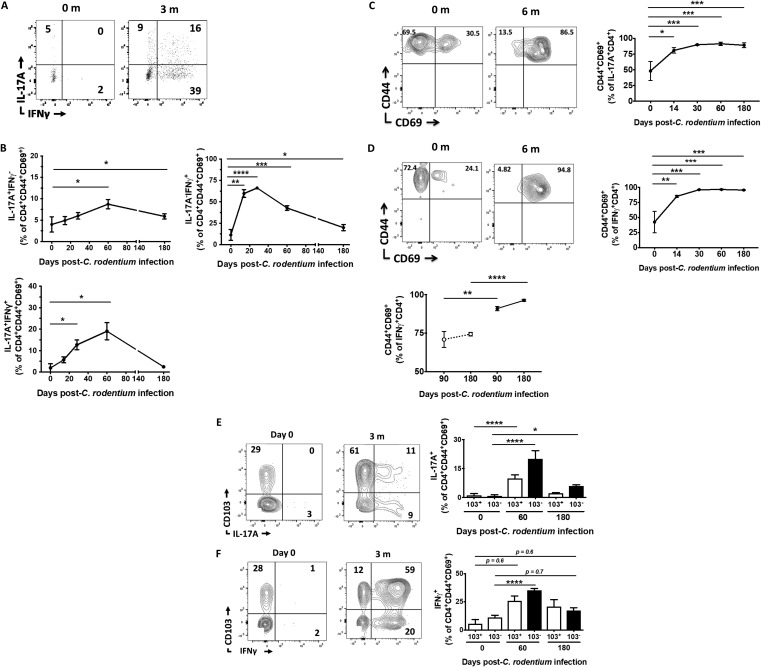
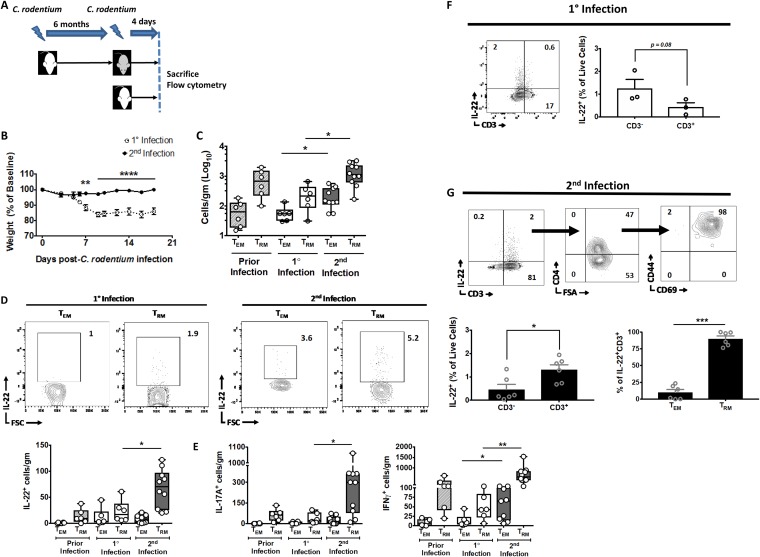
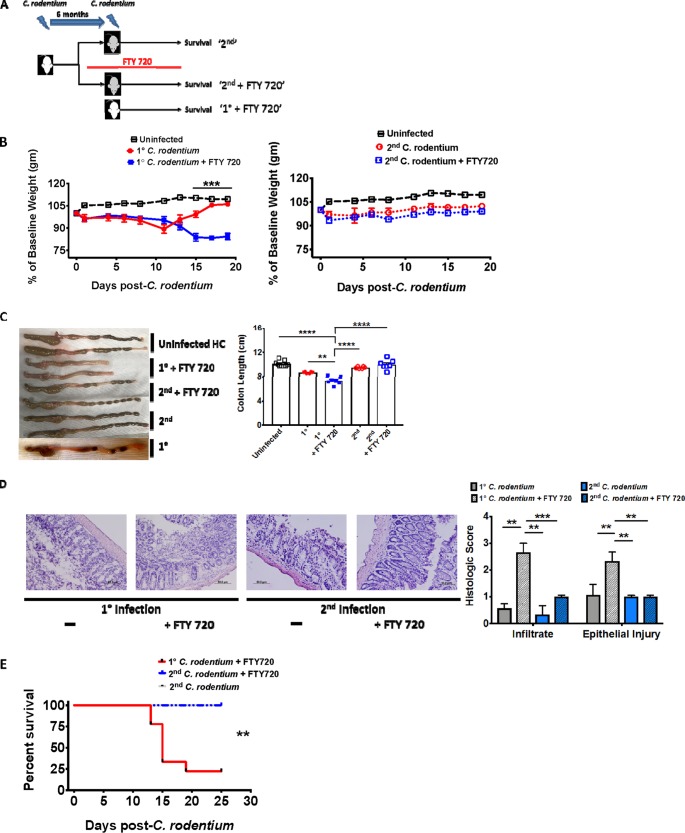
Tissue-resident memory T cells (TRM cells) are a novel population of tissue-restricted antigen-specific T cells. TRM cells are induced by pathogens and promote host defense against secondary infections. Although TRM cells cannot be detected in circulation, they are the major memory CD4+ and CD8+ T-cell population in tissues in mice and humans. Murine models of CD8+ TRM cells have shown that CD8+ TRM cells maintain tissue residency via CD69 and though tumor growth factor β-dependent induction of CD103. In contrast to CD8+ TRM cells, there are few models of CD4+ TRM cells. Thus, much less is known about the factors regulating the induction, maintenance, and host defense functions of CD4+ TRM cells. Citrobacter rodentium is known to induce IL-17+ and IL-22+ CD4+ T cells (Th17 and Th22 cells, respectively). Moreover, data from IL-22 reporter mice show that most IL-22+ cells in the colon 3 months after C. rodentium infection are CD4+ T cells. This collectively suggests that C. rodentium may induce CD4+ TRM cells. Here, we demonstrate that C. rodentium induces a population of IL-17A+ CD4+ T cells that are tissue restricted and antigen specific, thus meeting the criteria of CD4+ TRM cells. These cells expand and are a major source of IL-22 during secondary C. rodentium infection, even before the T-cell phase of the host response in primary infection. Finally, using FTY 720, which depletes circulating naive and effector T cells but not tissue-restricted T cells, we show that these CD4+ TRM cells can promote host defense.
Keywords: CD4+; Citrobacter; T cells; colitis; tissue-resident memory.
Copyright © 2019 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
