Hippo Signaling Controls NLR Family Pyrin Domain Containing 3 Activation and Governs Immunoregulation of Mesenchymal Stem Cells in Mouse Liver Injury
- PMID: 31063235
- PMCID: PMC6819196
- DOI: 10.1002/hep.30700
Hippo Signaling Controls NLR Family Pyrin Domain Containing 3 Activation and Governs Immunoregulation of Mesenchymal Stem Cells in Mouse Liver Injury
Abstract
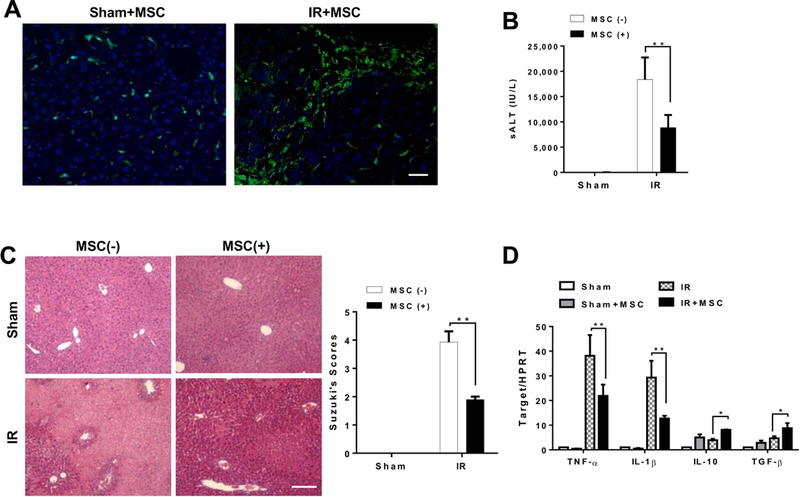
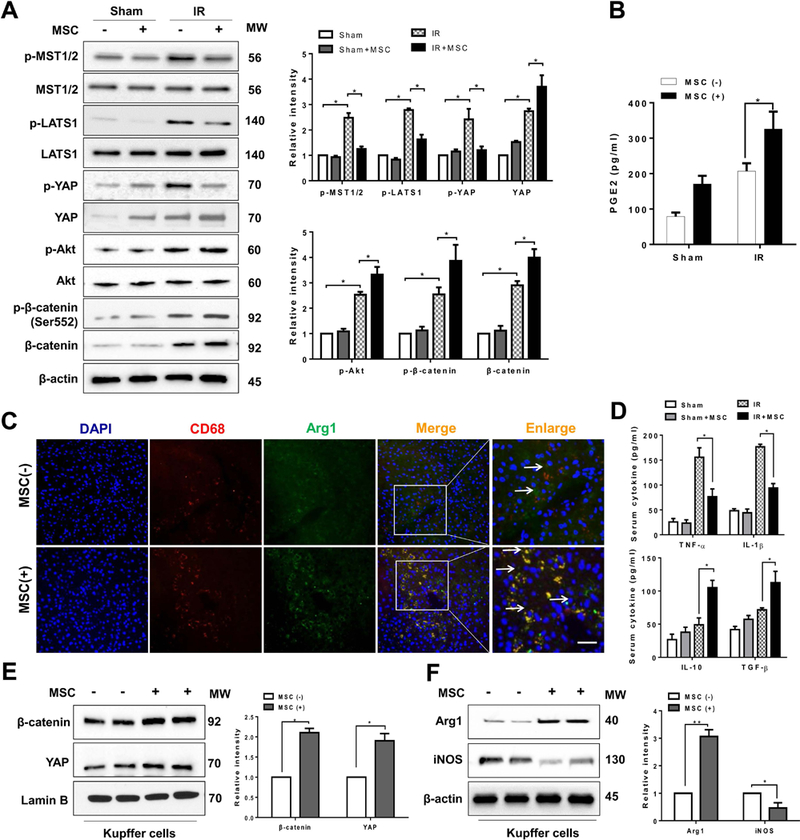
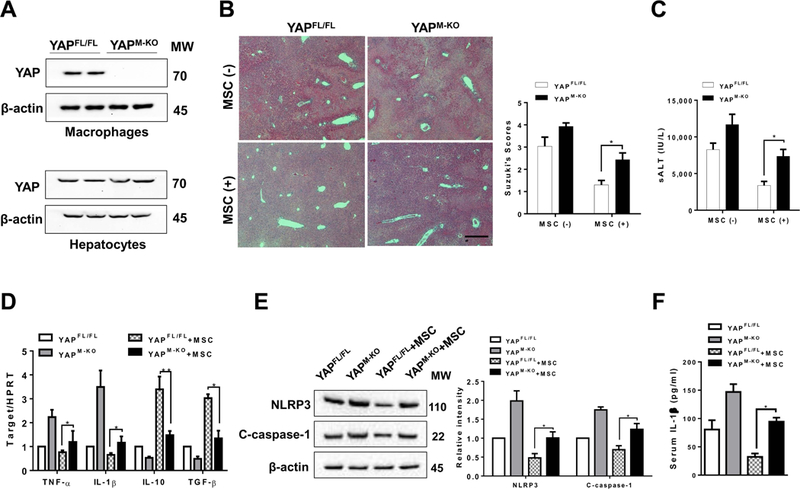
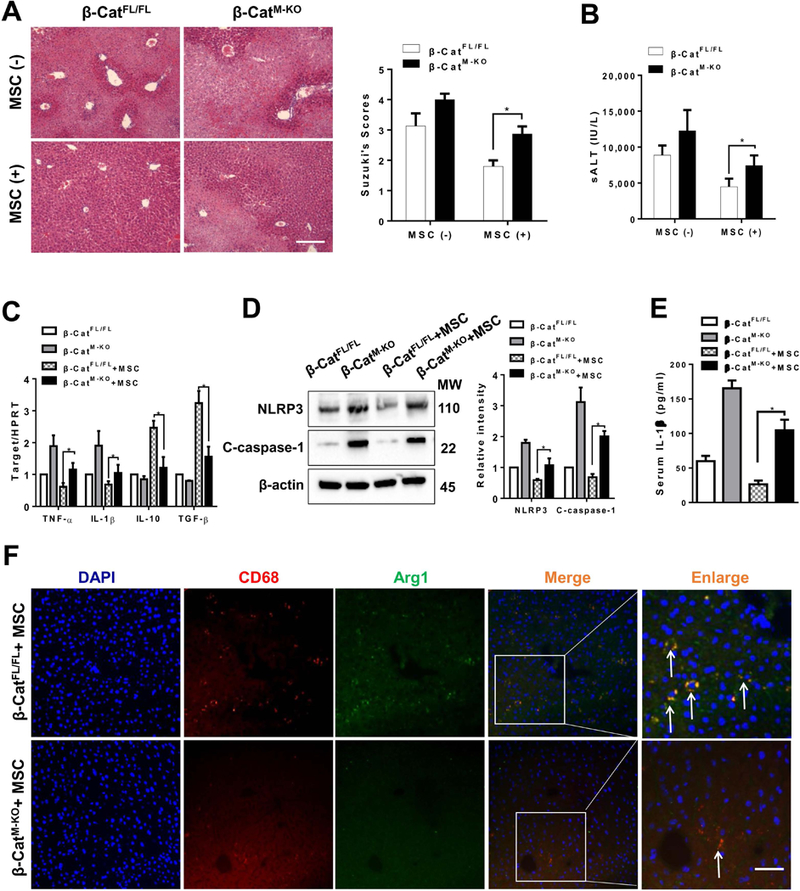
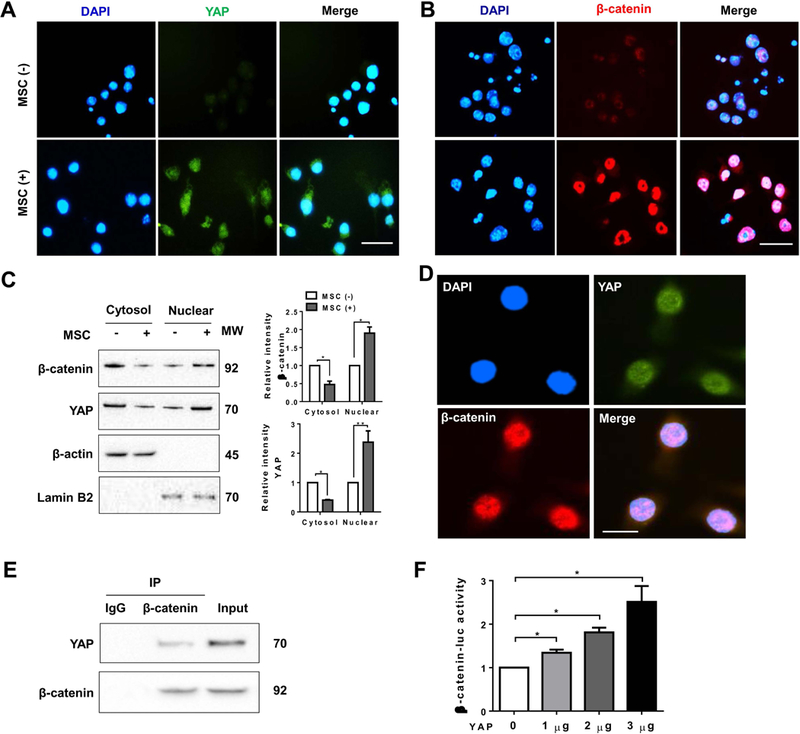
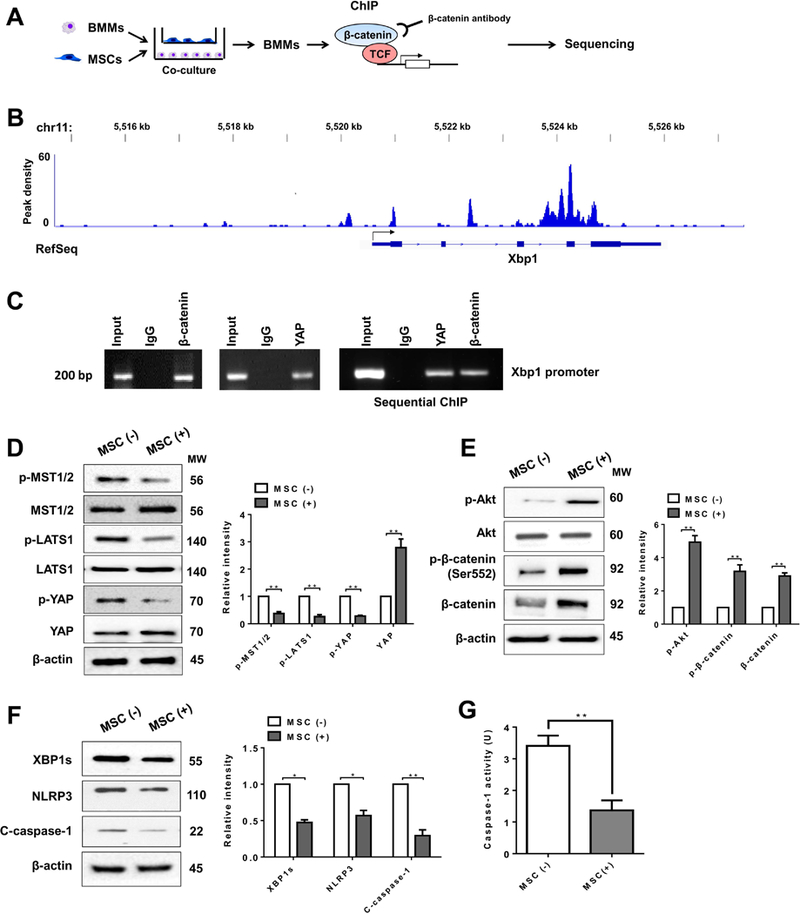
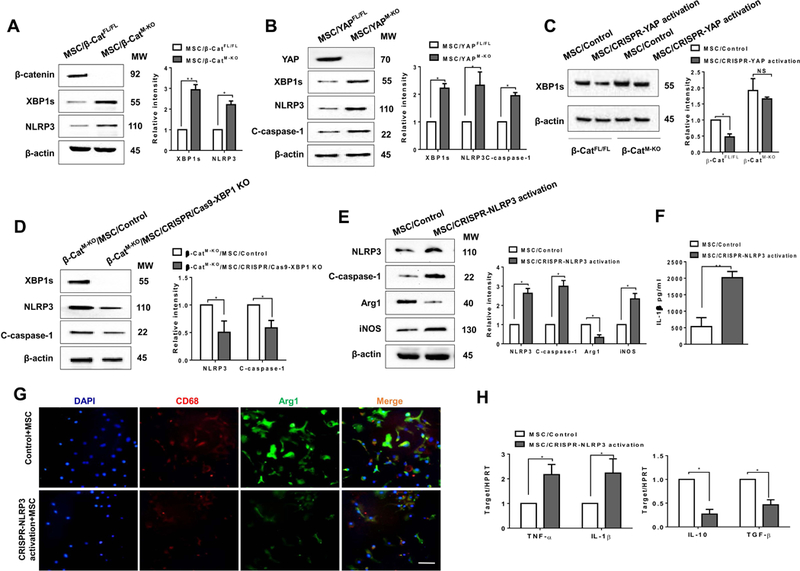
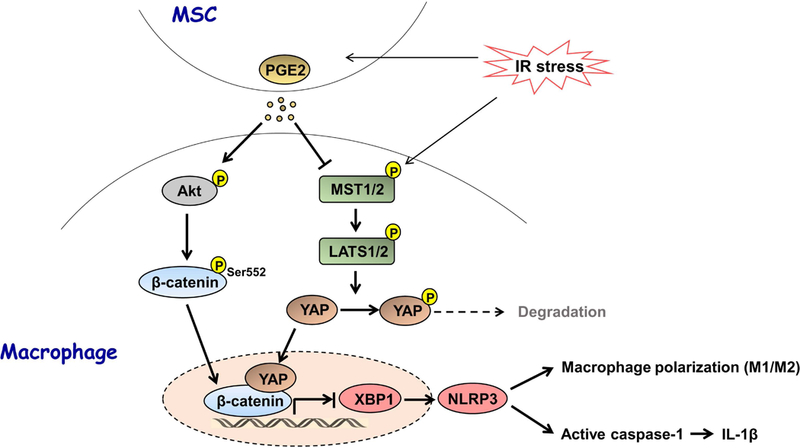
The Hippo pathway, an evolutionarily conserved protein kinase cascade, tightly regulates cell growth and survival. Activation of yes-associated protein (YAP), a downstream effector of the Hippo pathway, has been shown to modulate tissue inflammation. However, it remains unknown as to whether and how the Hippo-YAP signaling may control NLR family pyrin domain containing 3 (NLRP3) activation in mesenchymal stem cell (MSC)-mediated immune regulation during liver inflammation. In a mouse model of ischemia/reperfusion (IR)-induced liver sterile inflammatory injury, we found that adoptive transfer of MSCs reduced hepatocellular damage, shifted macrophage polarization from M1 to M2 phenotype, and diminished inflammatory mediators. MSC treatment reduced mammalian Ste20-like kinase 1/2 and large tumor suppressor 1 phosphorylation but augmented YAP and β-catenin expression with increased prostaglandin E2 production in ischemic livers. However, disruption of myeloid YAP or β-catenin in MSC-transferred mice exacerbated IR-triggered liver inflammation, enhanced NLRP3/caspase-1 activity, and reduced M2 macrophage phenotype. Using MSC/macrophage coculture system, we found that MSCs increased macrophage YAP and β-catenin nuclear translocation. Importantly, YAP and β-catenin colocalize in the nucleus while YAP interacts with β-catenin and regulates its target gene X-box binding protein 1 (XBP1), leading to reduced NLRP3/caspase-1 activity after coculture. Moreover, macrophage YAP or β-catenin deficiency augmented XBP1/NLRP3 while XBP1 deletion diminished NLRP3/caspase-1 activity. Increasing NLRP3 expression reduced M2 macrophage arginase1 but augmented M1 macrophage inducible nitric oxide synthase expression accompanied by increased interleukin-1β release. Conclusion: MSCs promote macrophage Hippo pathway, which in turn controls NLRP3 activation through a direct interaction between YAP and β-catenin and regulates XBP1-mediated NLRP3 activation, leading to reprograming macrophage polarization toward an anti-inflammatory M2 phenotype. Moreover, YAP functions as a transcriptional coactivator of β-catenin in MSC-mediated immune regulation. Our findings suggest a therapeutic target in MSC-mediated immunotherapy of liver sterile inflammatory injury.
© 2019 by the American Association for the Study of Liver Diseases.
Figures