

Defining the dynamic chromatin landscape of mouse nephron progenitors
- PMID: 31064740
- PMCID: PMC6550063
- DOI: 10.1242/bio.042754
Defining the dynamic chromatin landscape of mouse nephron progenitors
Abstract
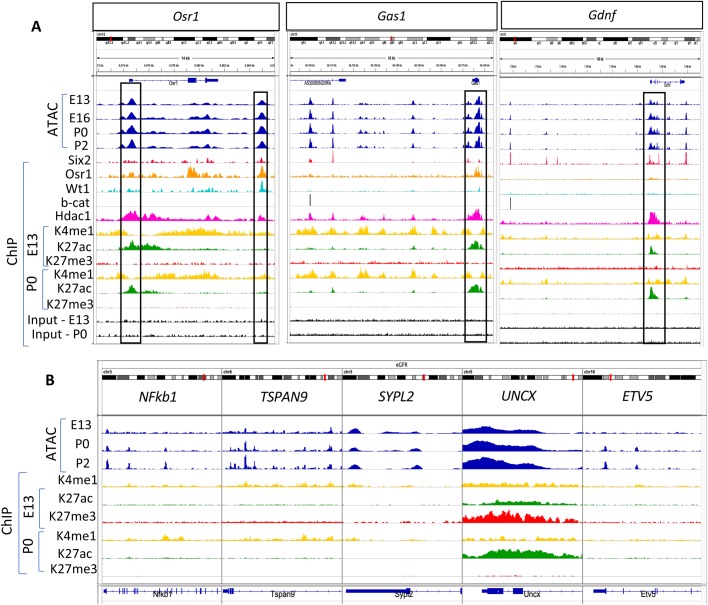
Six2+ cap mesenchyme cells, also called nephron progenitor cells (NPC), are precursors of all epithelial cell types of the nephron, the filtering unit of the kidney. Current evidence indicates that perinatal 'old' NPC have a greater tendency to exit the progenitor niche and differentiate into nascent nephrons than their embryonic 'young' counterpart. Understanding the underpinnings of NPC development may offer insights to rejuvenate old NPC and expand the progenitor pool. Here, we compared the chromatin landscape of young and old NPC and found common features reflecting their shared lineage but also intrinsic differences in chromatin accessibility and enhancer landscape supporting the view that old NPC are epigenetically poised for differentiation. Annotation of open chromatin regions and active enhancers uncovered the transcription factor Bach2 as a potential link between the pro-renewal MAPK/AP1 and pro-differentiation Six2/b-catenin pathways that might be of critical importance in regulation of NPC fate. Our data provide the first glimpse of the dynamic chromatin landscape of NPC and serve as a platform for future studies of the impact of genetic or environmental perturbations on the epigenome of NPC.
Keywords: ATAC-seq; ChIP-seq; Epigenetics; Kidney; Nephrogenesis.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures







References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
