

Preexisting Commensal Dysbiosis Is a Host-Intrinsic Regulator of Tissue Inflammation and Tumor Cell Dissemination in Hormone Receptor-Positive Breast Cancer
- PMID: 31064848
- PMCID: PMC6983951
- DOI: 10.1158/0008-5472.CAN-18-3464
Preexisting Commensal Dysbiosis Is a Host-Intrinsic Regulator of Tissue Inflammation and Tumor Cell Dissemination in Hormone Receptor-Positive Breast Cancer
Abstract
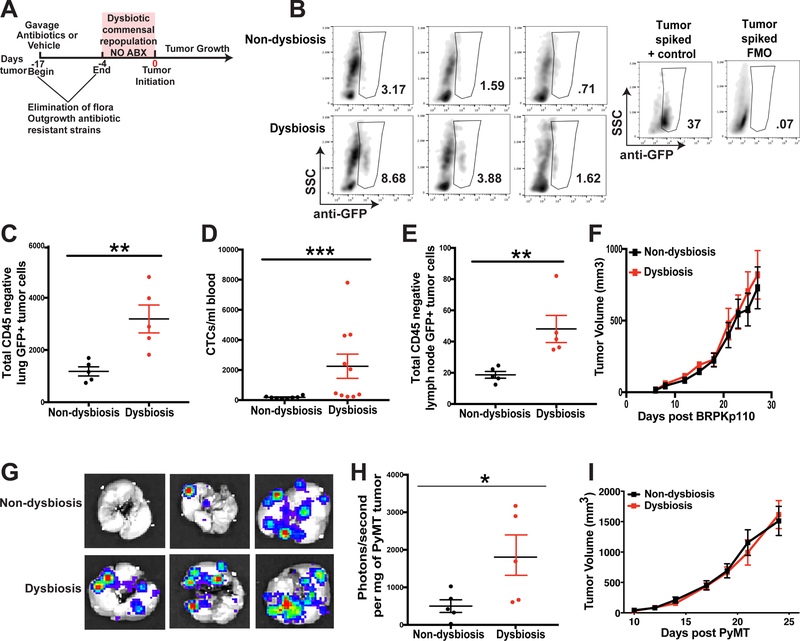
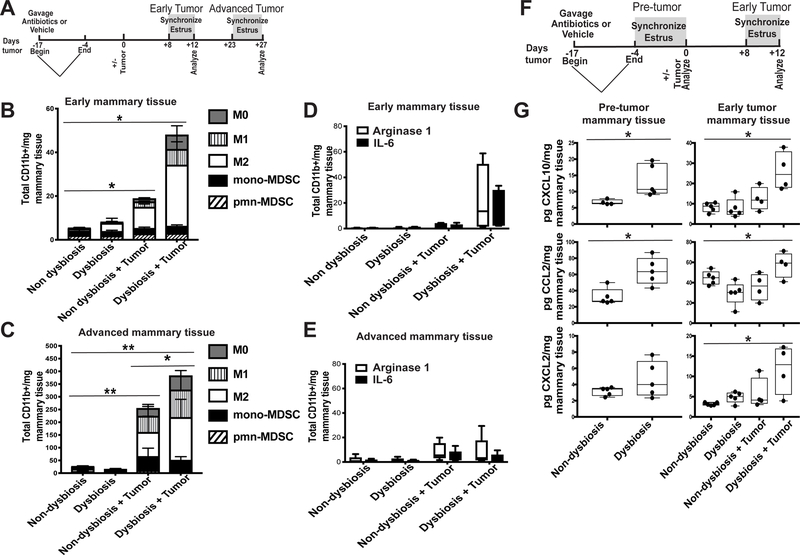
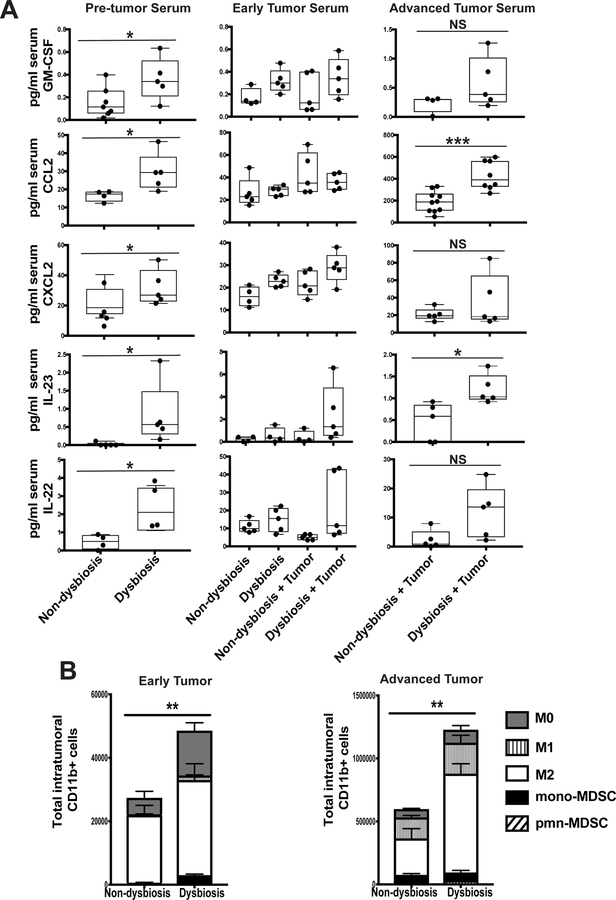
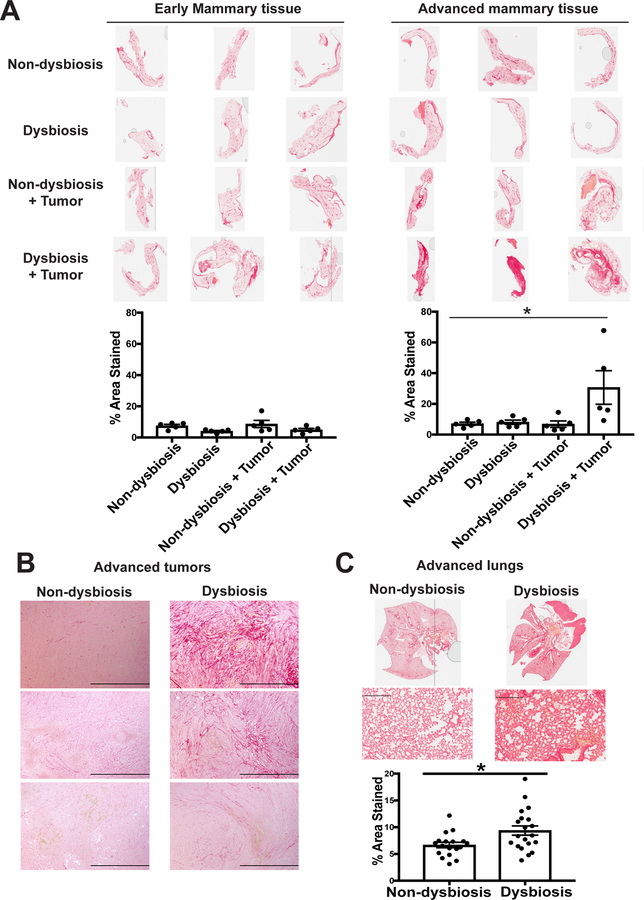
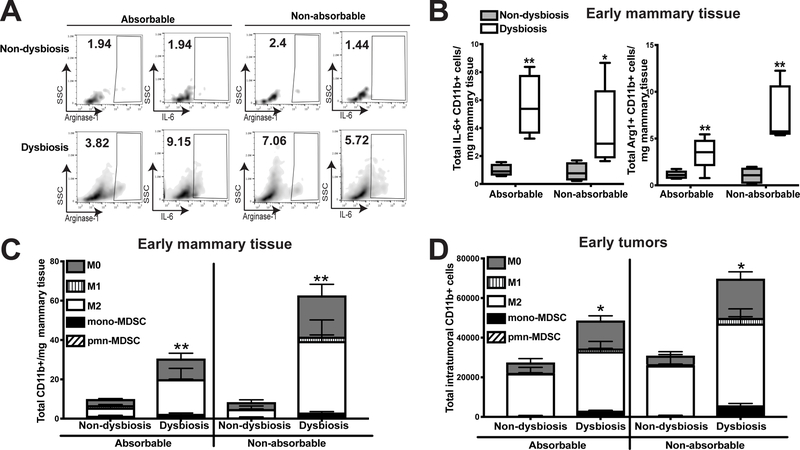
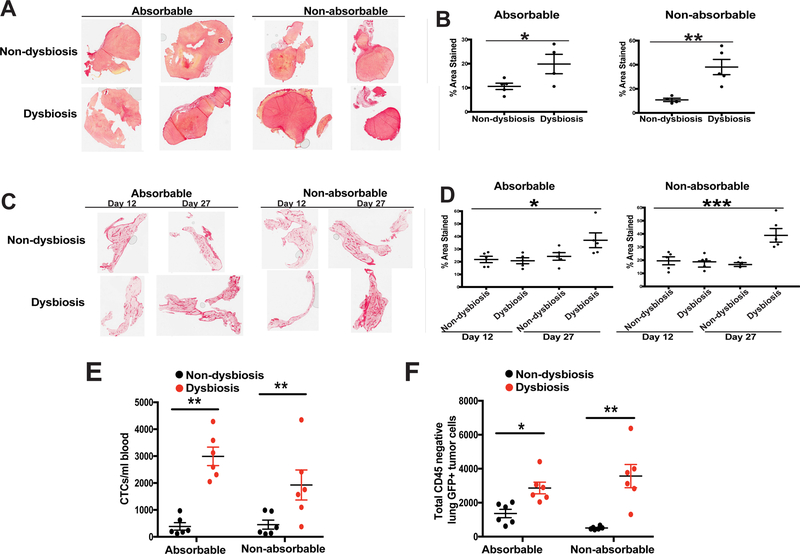
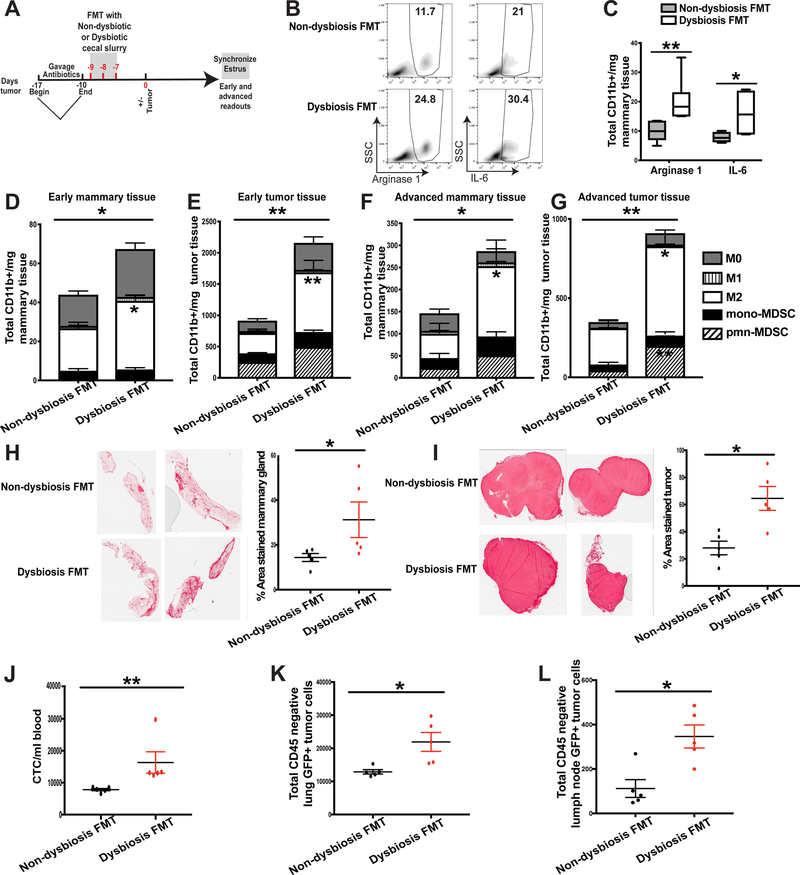
It is unknown why some patients with hormone receptor-positive (HR+) breast cancer present with more aggressive and invasive disease. Metastatic dissemination occurs early in disease and is facilitated by cross-talk between the tumor and tissue environment, suggesting that undefined host-intrinsic factors enhance early dissemination and the probability of developing metastatic disease. Here, we have identified commensal dysbiosis as a host-intrinsic factor associated with metastatic dissemination. Using a mouse model of HR+ mammary cancer, we demonstrate that a preestablished disruption of commensal homeostasis results in enhanced circulating tumor cells and subsequent dissemination to the tumor-draining lymph nodes and lungs. Commensal dysbiosis promoted early inflammation within the mammary gland that was sustained during HR+ mammary tumor progression. Furthermore, dysbiosis enhanced fibrosis and collagen deposition both systemically and locally within the tumor microenvironment and induced significant myeloid infiltration into the mammary gland and breast tumor. These effects were recapitulated both by directly targeting gut microbes using nonabsorbable antibiotics and by fecal microbiota transplantation of dysbiotic cecal contents, demonstrating the direct impact of gut dysbiosis on mammary tumor dissemination. This study identifies dysbiosis as a preexisting, host-intrinsic regulator of tissue inflammation, myeloid recruitment, fibrosis, and dissemination of tumor cells in HR+ breast cancer. SIGNIFICANCE: Identification of commensal dysbiosis as a host-intrinsic factor mediating evolution of metastatic breast cancer allows for development of interventions or diagnostic tools for patients at highest risk for developing metastatic disease.See related commentary by Ingman, p. 3539.
©2019 American Association for Cancer Research.
Conflict of interest statement
Figures
Comment in
-
The Gut Microbiome: A New Player in Breast Cancer Metastasis.Cancer Res. 2019 Jul 15;79(14):3539-3541. doi: 10.1158/0008-5472.CAN-19-1698. Cancer Res. 2019. PMID: 31308136
References
-
- Burstein HJ, Cirrincione CT, Barry WT, Chew HK, Tolaney SM, Lake DE, et al. Endocrine therapy with or without inhibition of epidermal growth factor receptor and human epidermal growth factor receptor 2: a randomized, double-blind, placebo-controlled phase III trial of fulvestrant with or without lapatinib for postmenopausal women with hormone receptor-positive advanced breast cancer-CALGB 40302 (Alliance). J Clin Oncol 2014;32:3959–66. 10.1200/JCO.2014.56.7941. Epub 2014 Oct 27. - DOI - PMC - PubMed
-
- Song F, Zhang J, Li S, Wu J, Jin T, Qin J, et al. ER-positive breast cancer patients with more than three positive nodes or grade 3 tumors are at high risk of late recurrence after 5-year adjuvant endocrine therapy. Onco Targets Ther 2017;10:4859–4867.: 10.2147/OTT.S142698. eCollection 2017. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
