

Evolutionary comparison of competitive protein-complex formation of MYB, bHLH, and WDR proteins in plants
- PMID: 31071215
- PMCID: PMC6598095
- DOI: 10.1093/jxb/erz155
Evolutionary comparison of competitive protein-complex formation of MYB, bHLH, and WDR proteins in plants
Abstract
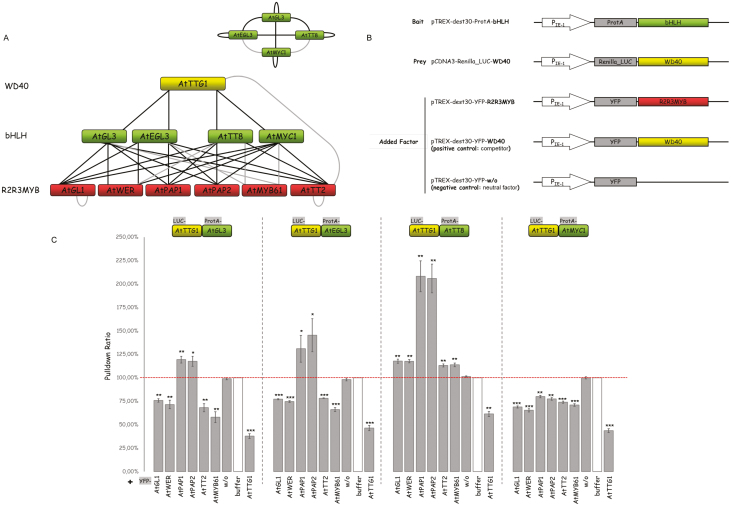
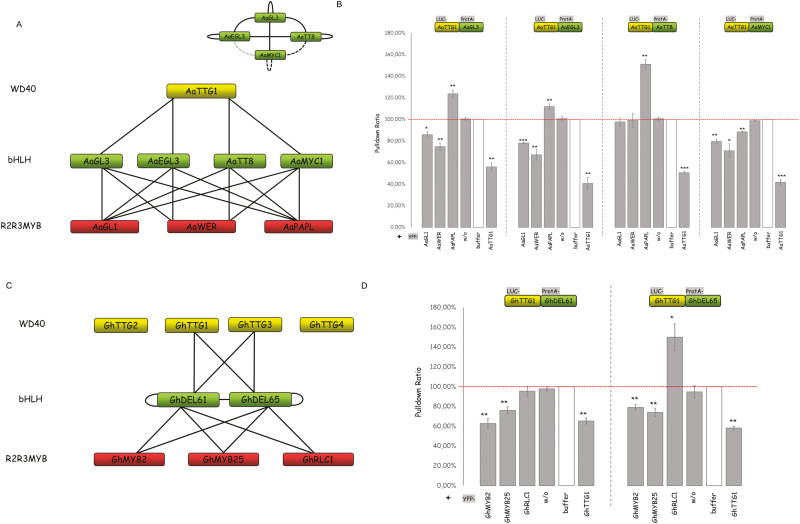
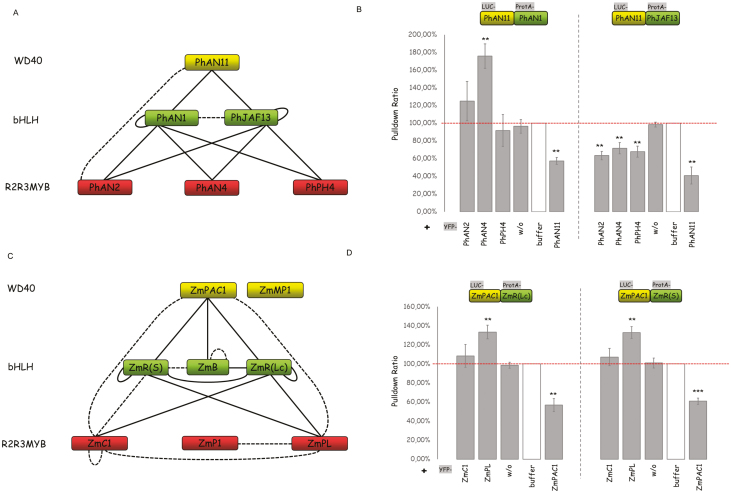
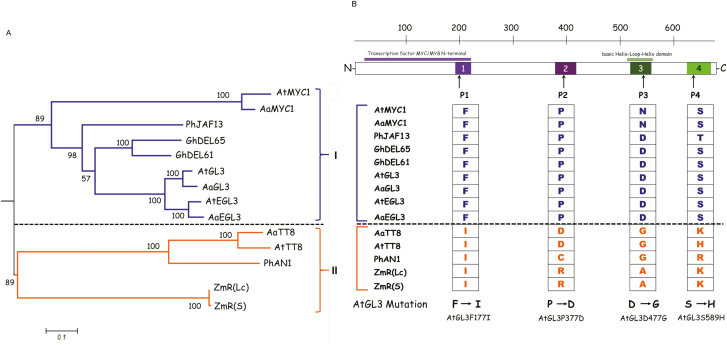
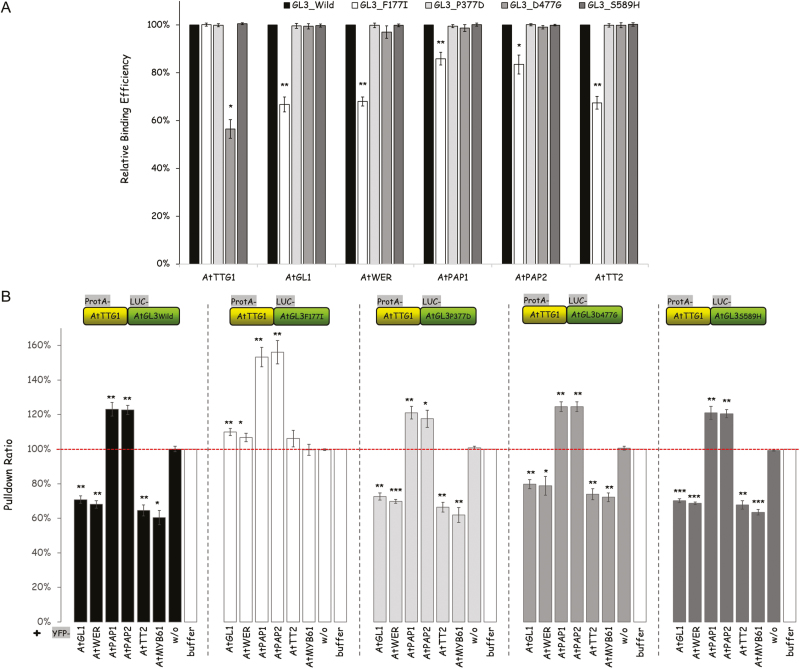
A protein complex consisting of a MYB, basic Helix-Loop-Helix, and a WDR protein, the MBW complex, regulates five traits, namely the production of anthocyanidin, proanthocyanidin, and seed-coat mucilage, and the development of trichomes and root hairs. For complexes involved in trichome and root hair development it has been shown that the interaction of two MBW proteins can be counteracted by the respective third protein (called competitive complex formation). We examined competitive complex formation for selected MBW proteins from Arabidopsis thaliana, Arabis alpina, Gossypium hirsutum, Petunia hybrida, and Zea mays. Quantitative analyses of the competitive binding of MYBs and WDRs to bHLHs were done by pull-down assays using ProtA- and luciferase-tagged proteins expressed in human HEC cells. We found that some bHLHs show competitive complex formation whilst others do not. Competitive complex formation strongly correlated with a phylogenetic tree constructed with the bHLH proteins under investigation, suggesting a functional relevance. We demonstrate that this different behavior can be explained by changes in one amino acid and that this position is functionally relevant in trichome development but not in anthocyanidin regulation.
Keywords: Arabidopsis thaliana; Arabis alpina; Gossypium hirsutum; Petunia hybrida; Zea mays; MBW complex; competitive complex formation; evolution.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures

References
-
- Albert NW, Lewis DH, Zhang H, Schwinn KE, Jameson PE, Davies KM. 2011. Members of an R2R3-MYB transcription factor family in Petunia are developmentally and environmentally regulated to control complex floral and vegetative pigmentation patterning. The Plant Journal 65, 771–784. - PubMed
-
- Altschul SF, Gish W. 1996. Local alignment statistics. Methods in Enzymology 266, 460–480. - PubMed
-
- Bailey TL, Gribskov M. 1998. Combining evidence using p-values: application to sequence homology searches. Bioinformatics 14, 48–54. - PubMed
-
- Baudry A, Caboche M, Lepiniec L. 2006. TT8 controls its own expression in a feedback regulation involving TTG1 and homologous MYB and bHLH factors, allowing a strong and cell-specific accumulation of flavonoids in Arabidopsis thaliana. Plant Journal 46, 768–779. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
