

The Catalytic Subunit β of PKA Affects Energy Balance and Catecholaminergic Activity
- PMID: 31073546
- PMCID: PMC6503631
- DOI: 10.1210/js.2019-00029
The Catalytic Subunit β of PKA Affects Energy Balance and Catecholaminergic Activity
Abstract
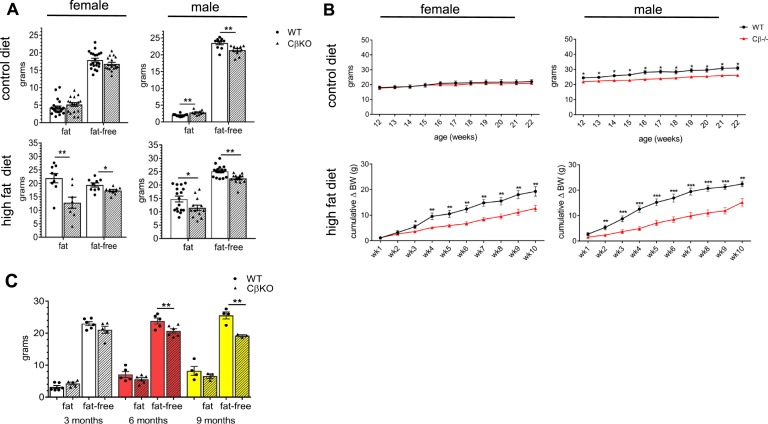
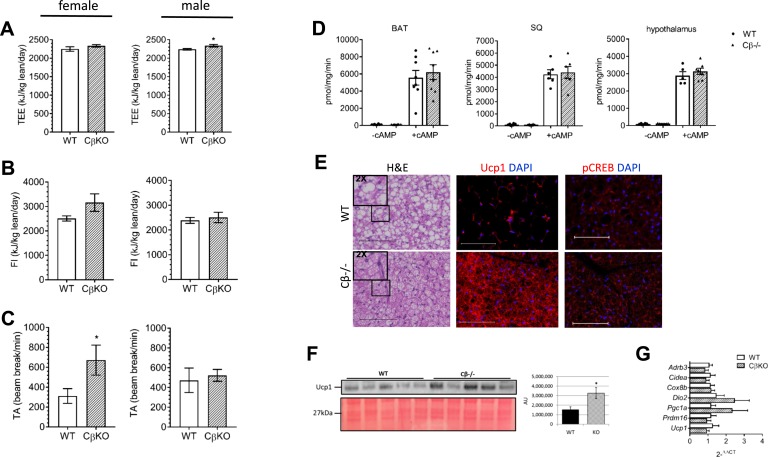
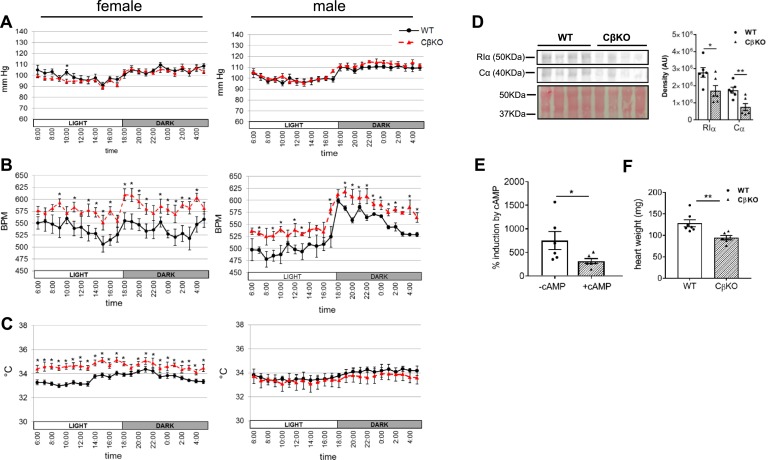
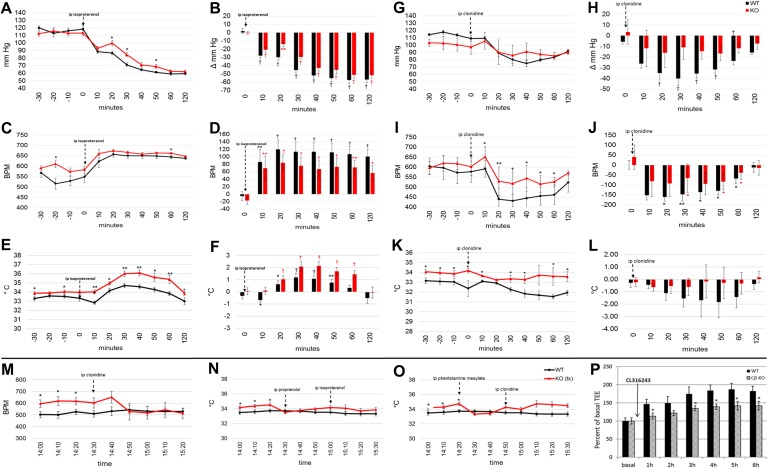
The protein kinase A (PKA) signaling system mediates the effects of numerous hormones, neurotransmitters, and other molecules to regulate metabolism, cardiac function, and more. PKA defects may lead to diverse phenotypes that largely depend on the unique expression profile of the affected subunit. Deletion of the Prkarcb gene, which codes for PKA catalytic subunit β (Cβ), protects against diet-induced obesity (DIO), yet the mechanism for this phenotype remains unclear. We hypothesized that metabolic rate would be increased in Cβ knockout (KO) mice, which could explain DIO resistance. Male, but not female, CβKO mice had increased energy expenditure, and female but not male CβKO mice had increased subcutaneous temperature and increased locomotor activity compared with wild-type (WT) littermates. Urinary norepinephrine (NE) and normetanephrine were elevated in female CβKO mice. CβKO mice had increased heart rate (HR); blocking central NE release normalized HR to that of untreated WT mice. Basal and stimulated PKA enzymatic activities were unchanged in adipose tissue and heart and varied in different brain regions, suggesting that Prkacb deletion may mediate signaling changes in specific brain nuclei and may be less important in the peripheral regulation of PKA expression and activity. This is a demonstration of a distinct effect of the PKA Cβ catalytic subunit on catecholamines and sympathetic nerve signaling. The data provide an unexpected explanation for the metabolic phenotype of CβKO mice. Finally, the sexual dimorphism is consistent with mouse models of other PKA subunits and adds to the importance of these findings regarding the PKA system in human metabolism.
Keywords: PKA; cardiovascular; catecholamines; energy balance; sympathetic outflow.
Figures


References
-
- Cummings DE, Brandon EP, Planas JV, Motamed K, Idzerda RL, McKnight GS. Genetically lean mice result from targeted disruption of the RII beta subunit of protein kinase A. Nature. 1996;382(6592):622–626. - PubMed
-
- London E, Nesterova M, Sinaii N, Szarek E, Chanturiya T, Mastroyannis SA, Gavrilova O, Stratakis CA. Differentially regulated protein kinase A (PKA) activity in adipose tissue and liver is associated with resistance to diet-induced obesity and glucose intolerance in mice that lack PKA regulatory subunit type IIα. Endocrinology. 2014;155(9):3397–3408. - PMC - PubMed
-
- Enns LC, Morton JF, Mangalindan RS, McKnight GS, Schwartz MW, Kaeberlein MR, Kennedy BK, Rabinovitch PS, Ladiges WC. Attenuation of age-related metabolic dysfunction in mice with a targeted disruption of the Cbeta subunit of protein kinase A. J Gerontol A Biol Sci Med Sci. 2009;64(12):1221–1231. - PMC - PubMed
-
- Planas JV, Cummings DE, Idzerda RL, McKnight GS. Mutation of the RIIbeta subunit of protein kinase A differentially affects lipolysis but not gene induction in white adipose tissue. J Biol Chem. 1999;274(51):36281–36287. - PubMed
-
- Schreyer SA, Cummings DE, McKnight GS, LeBoeuf RC. Mutation of the RIIbeta subunit of protein kinase A prevents diet-induced insulin resistance and dyslipidemia in mice. Diabetes. 2001;50(11):2555–2562. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials