

Structural, Kinetic, and Mechanistic Analysis of an Asymmetric 4-Oxalocrotonate Tautomerase Trimer
- PMID: 31074977
- PMCID: PMC6948721
- DOI: 10.1021/acs.biochem.9b00303
Structural, Kinetic, and Mechanistic Analysis of an Asymmetric 4-Oxalocrotonate Tautomerase Trimer
Abstract
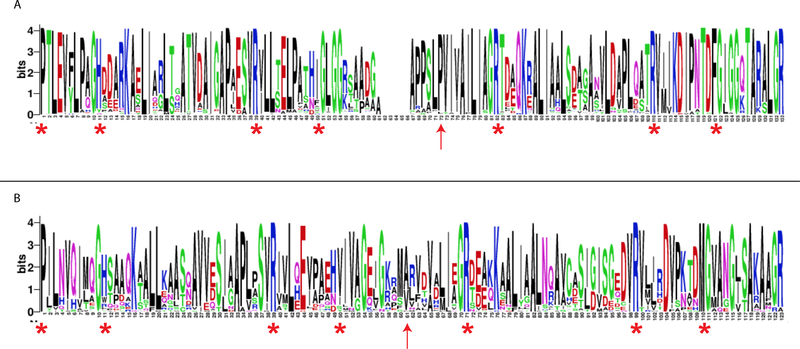
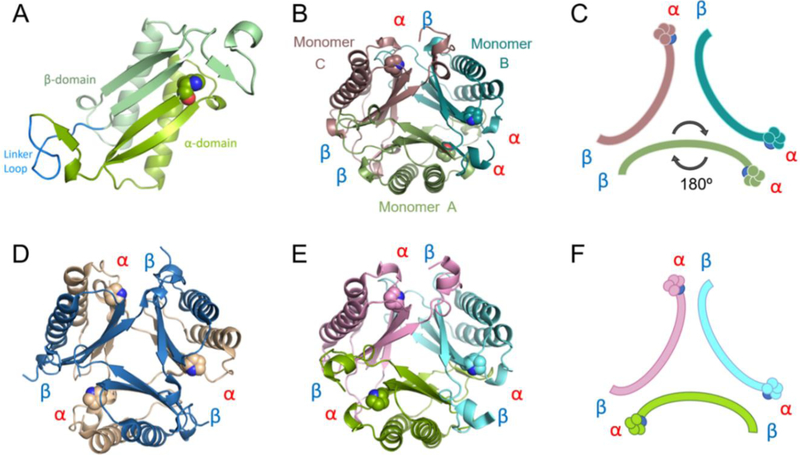
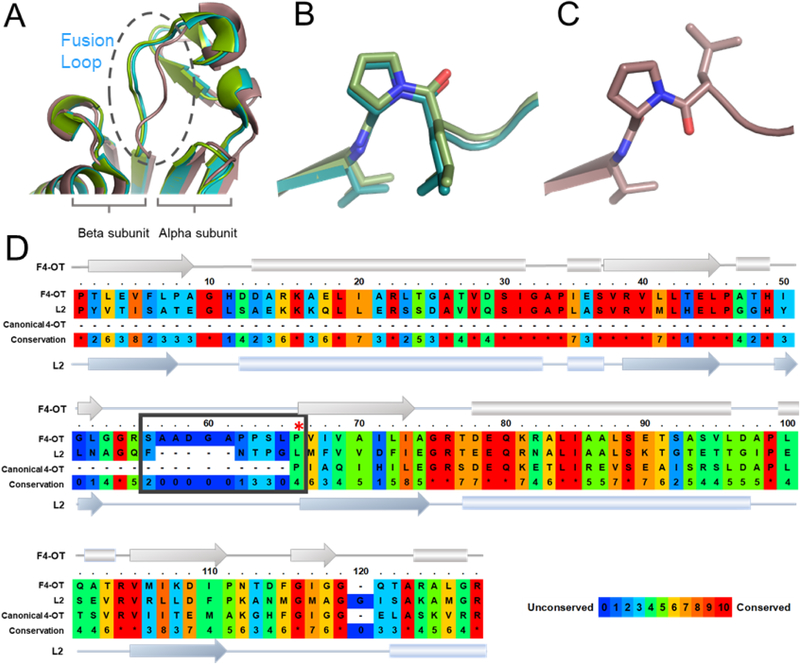
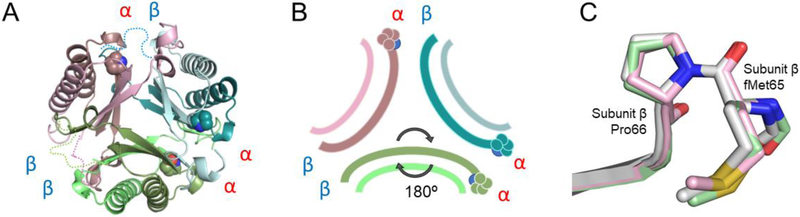
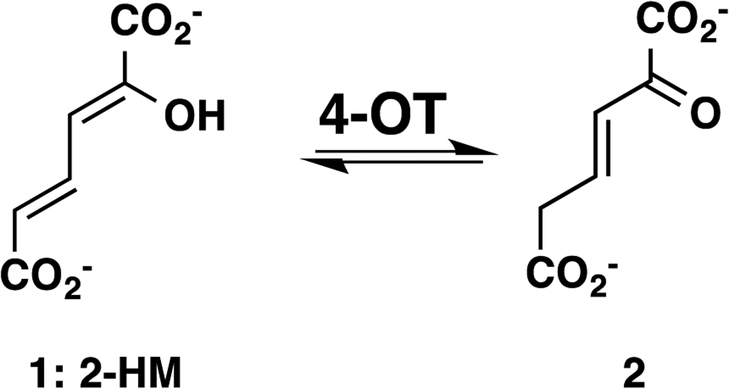
A 4-oxalocrotonate tautomerase (4-OT) trimer has been isolated from Burkholderia lata, and a kinetic, mechanistic, and structural analysis has been performed. The enzyme is the third described oligomer state for 4-OT along with a homo- and heterohexamer. The 4-OT trimer is part of a small subset of sequences (133 sequences) within the 4-OT subgroup of the tautomerase superfamily (TSF). The TSF has two distinct features: members are composed of a single β-α-β unit (homo- and heterohexamer) or two consecutively joined β-α-β units (trimer) and generally have a catalytic amino-terminal proline. The enzyme, designated as fused 4-OT, functions as a 4-OT where the active site groups (Pro-1, Arg-39, Arg-76, Phe-115, Arg-127) mirror those in the canonical 4-OT from Pseudomonas putida mt-2. Inactivation by 2-oxo-3-pentynoate suggests that Pro-1 of fused 4-OT has a low p Ka enabling the prolyl nitrogen to function as a general base. A remarkable feature of the fused 4-OT is the absence of P3 rotational symmetry in the structure (1.5 Å resolution). The asymmetric arrangement of the trimer is not due to the fusion of the two β-α-β building blocks because an engineered "unfused" variant that breaks the covalent bond between the two units (to generate a heterohexamer) assumes the same asymmetric oligomerization state. It remains unknown how the different active site configurations contribute to the observed overall activities and whether the asymmetry has a biological purpose or role in the evolution of TSF members.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Whitman CP, Aird BA, Gillespie WR, and Stolowich NJ (1991) Chemical and enzymatic ketonization of 2-hydroxymuconate, a conjugated enol. J. Am. Chem. Soc 113, 3154–3162.
-
- Dagley S (1978) Pathways for the utilization of organic growth substrates, in The Bacteria: A Treatise on Structure and Function (Ornston LN and Sokatch JR, Eds), pp. 305–388, Academic Press, New York.
-
- Stivers JT, Abeygunawardana C, Mildvan AS, Hajipour G, Whitman CP, and Chen LH (1996) Catalytic role of the amino-terminal proline in 4-oxalocrotonate tautomerase: affinity labeling and heteronuclear NMR studies. Biochemistry 35, 803–813. - PubMed
-
- Stivers JT, Abeygunawardana, Mildvan AS, Hajipour G, and Whitman CP (1996) 4-Oxalocrotonate tautomerase: pH dependences of catalysis and pKa values of active site residues. Biochemistry 35, 814–823. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
