

APETALA2 control of barley internode elongation
- PMID: 31076487
- PMCID: PMC6589076
- DOI: 10.1242/dev.170373
APETALA2 control of barley internode elongation
Abstract
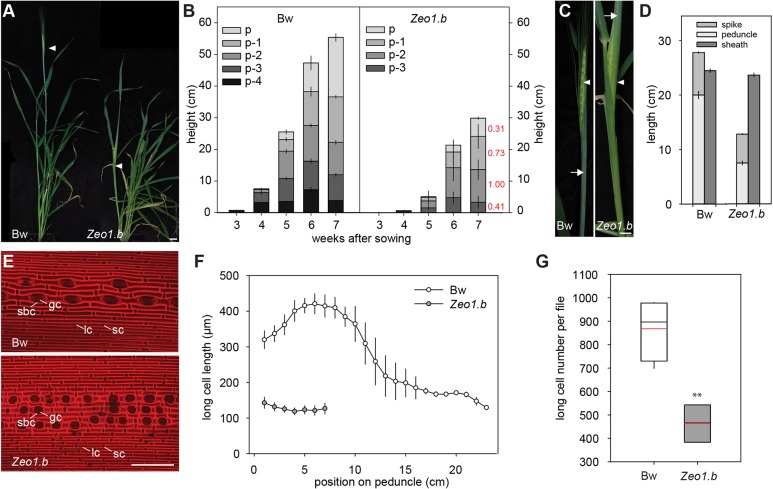
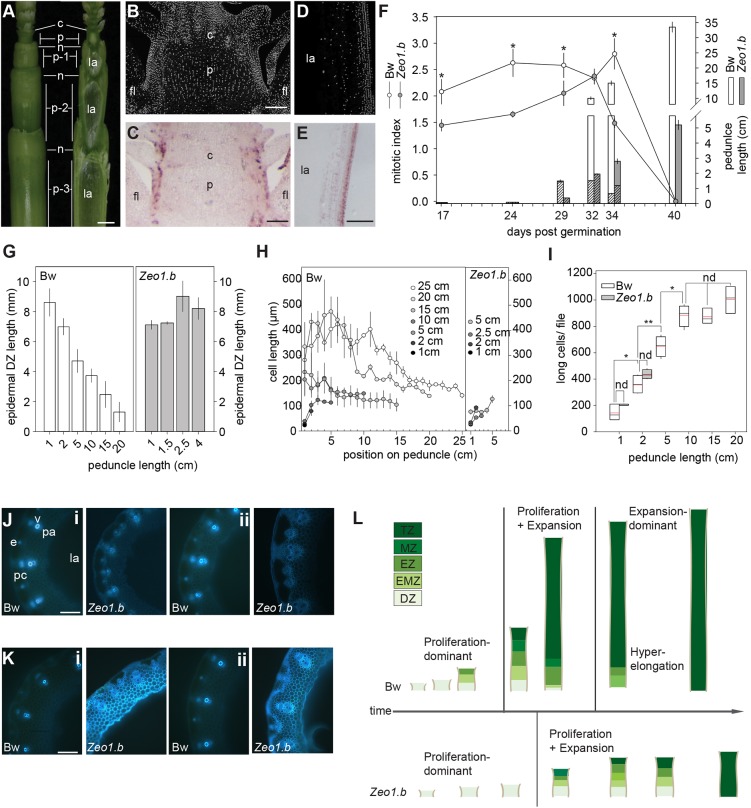
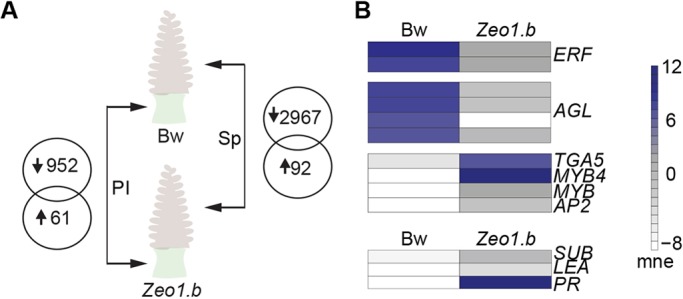
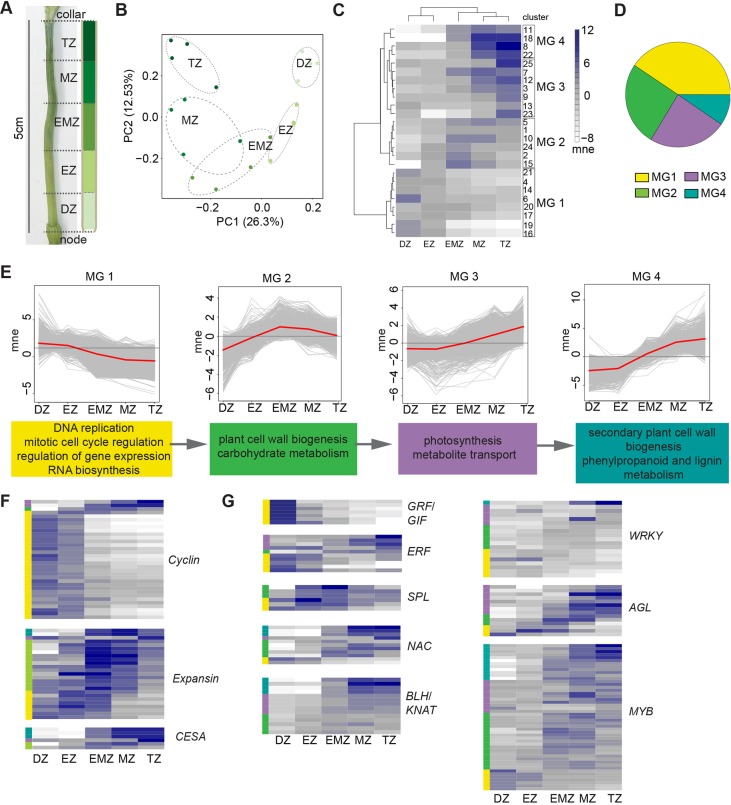
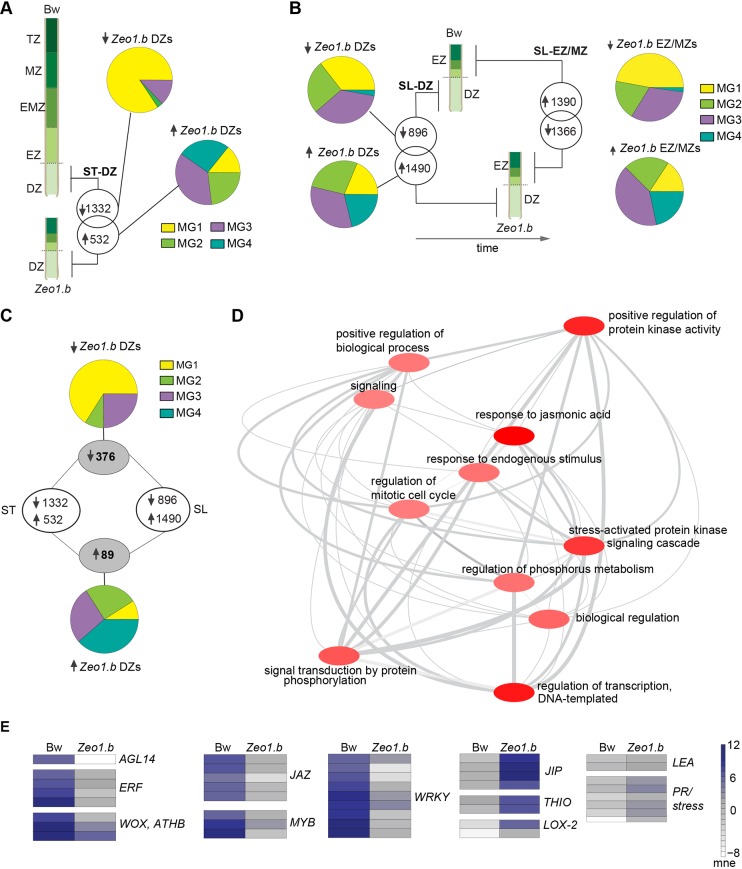
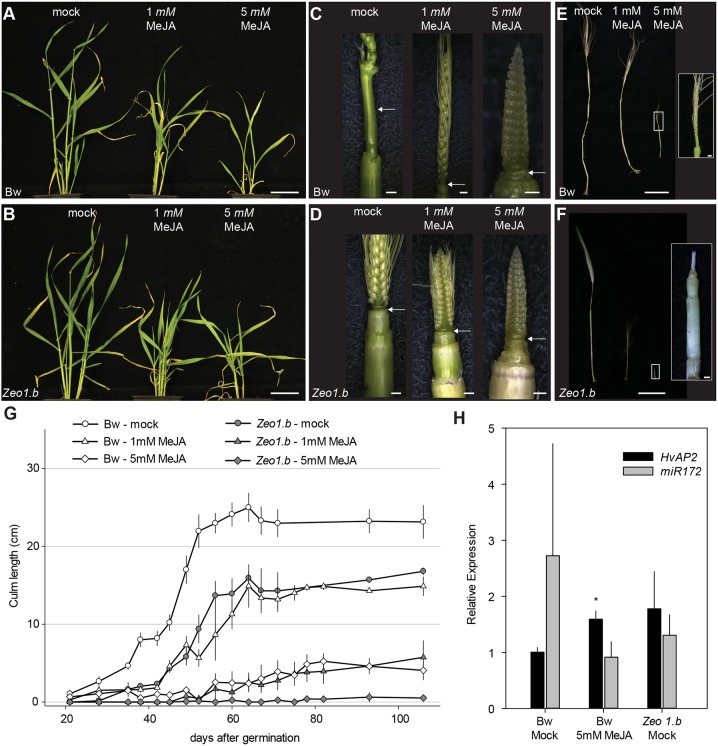
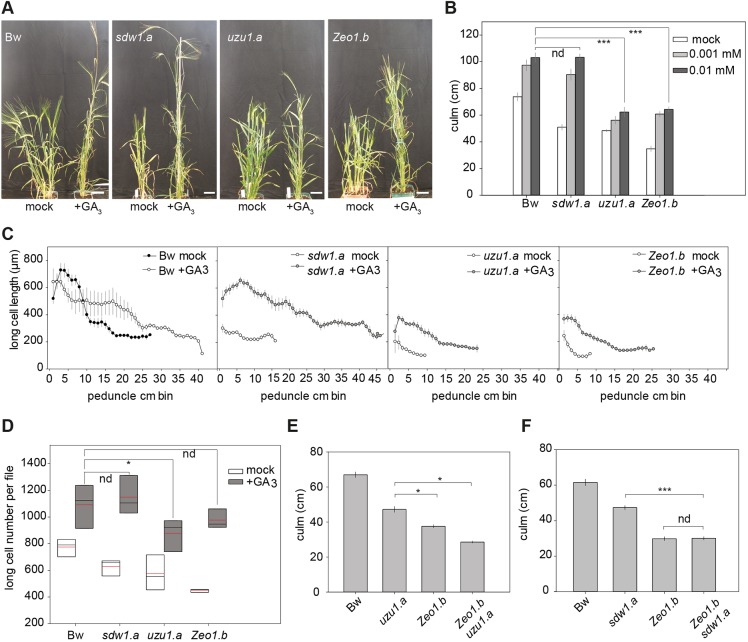
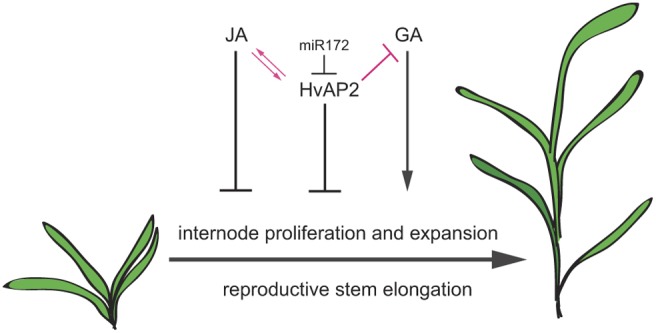
Many plants dramatically elongate their stems during flowering, yet how this response is coordinated with the reproductive phase is unclear. We demonstrate that microRNA (miRNA) control of APETALA2 (AP2) is required for rapid, complete elongation of stem internodes in barley, especially of the final 'peduncle' internode directly underneath the inflorescence. Disrupted miR172 targeting of AP2 in the Zeo1.b barley mutant caused lower mitotic activity, delayed growth dynamics and premature lignification in the peduncle leading to fewer and shorter cells. Stage- and tissue-specific comparative transcriptomics between Zeo1.b and its parent cultivar showed reduced expression of proliferation-associated genes, ectopic expression of maturation-related genes and persistent, elevated expression of genes associated with jasmonate and stress responses. We further show that applying methyl jasmonate (MeJA) phenocopied the stem elongation of Zeo1.b, and that Zeo1.b itself was hypersensitive to inhibition by MeJA but less responsive to promotion by gibberellin. Taken together, we propose that miR172-mediated restriction of AP2 may modulate the jasmonate pathway to facilitate gibberellin-promoted stem growth during flowering.
Keywords: APETALA2; Cereal development; Intercalary meristem; Jasmonate; Phase change; Stem elongation.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Baute J., Herman D., Coppens F., De Block J., Slabbinck B., Dell'Acqua M., Pè M. E., Maere S., Nelissen H. and Inze D. (2015). Correlation analysis of the transcriptome of growing leaves with mature leaf parameters in a maize RIL population. Genome Biol. 16, 168 10.1186/s13059-015-0735-9 - DOI - PMC - PubMed
-
- Bell A. D. and Bryan A. (2008). Plant Form: An Illustrated Guide to Flowering Plant Morphology. Timber Press.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
