

Seeing is believing: our evolving view of kinetochore structure, composition, and assembly
- PMID: 31078123
- PMCID: PMC6756984
- DOI: 10.1016/j.ceb.2019.03.016
Seeing is believing: our evolving view of kinetochore structure, composition, and assembly
Abstract
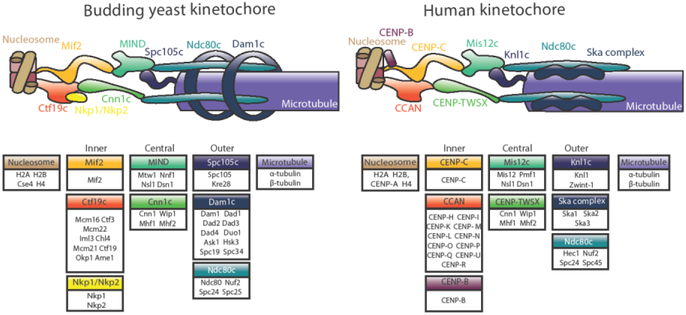
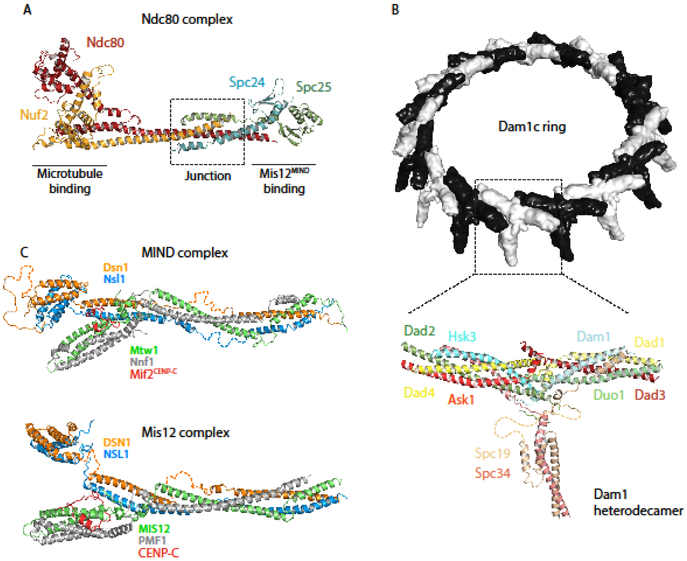
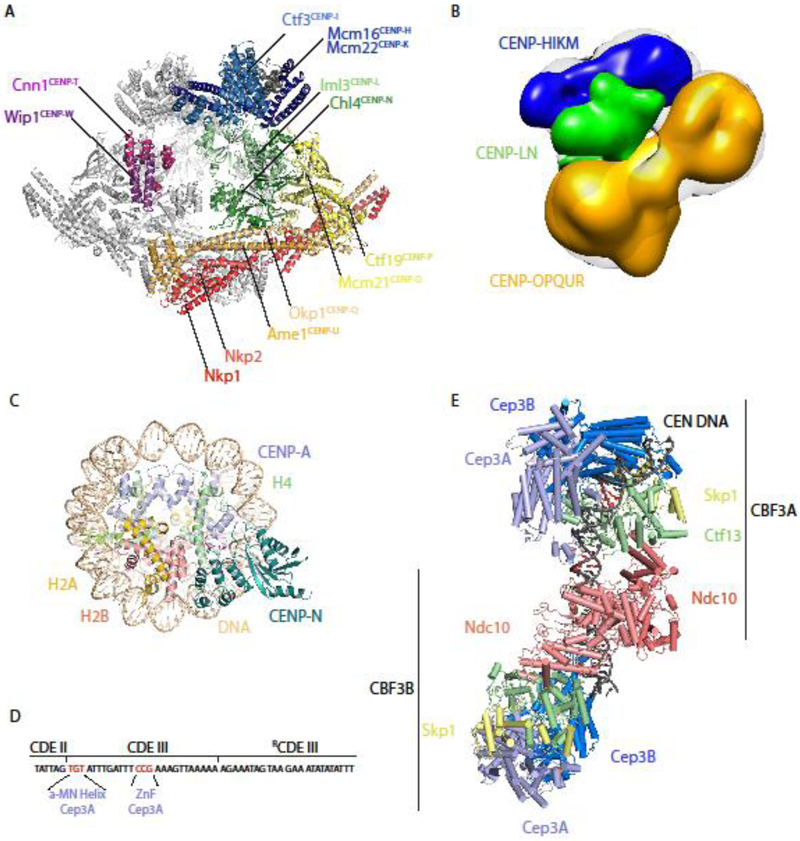
This review highlights three recent trends in the field of kinetochore biology: the proliferation of structural data for kinetochore protein complexes (including CBF3, Dam1c, Mis12cMIND, and CENP-NLChl4/Iml3); the growing consensus that the kinetochore is a dynamic structure whose composition changes as the cell cycle progresses; and the mounting evidence of multiple pathways whereby the microtubule-binding elements of the outer kinetochore may be recruited by inner kinetochore proteins. Our focus is on the two best-studied systems in the field: human and budding yeast kinetochores. This review will demonstrate the remarkable similarity of these two systems, as well as their intriguing differences.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
