

Mitochondrial Dysfunction Leads to Cortical Under-Connectivity and Cognitive Impairment
- PMID: 31079872
- PMCID: PMC6668992
- DOI: 10.1016/j.neuron.2019.04.013
Mitochondrial Dysfunction Leads to Cortical Under-Connectivity and Cognitive Impairment
Abstract
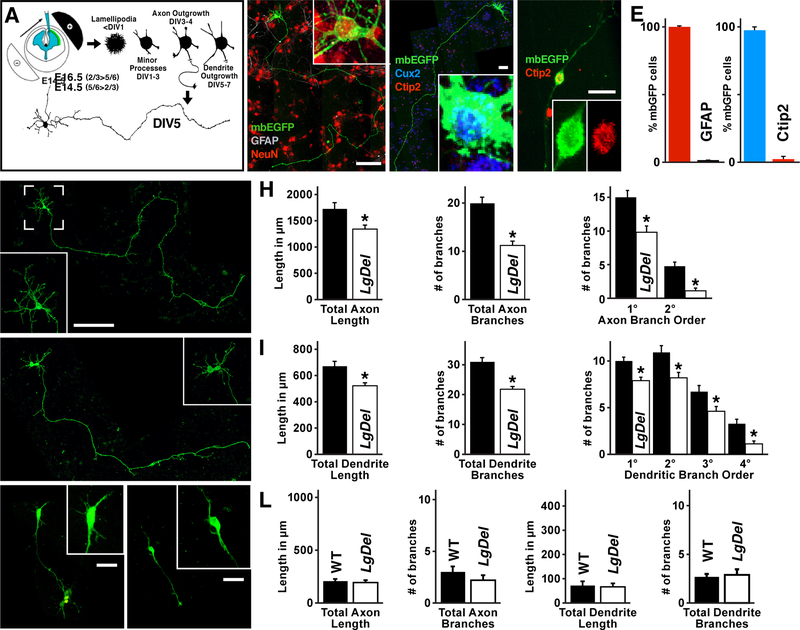
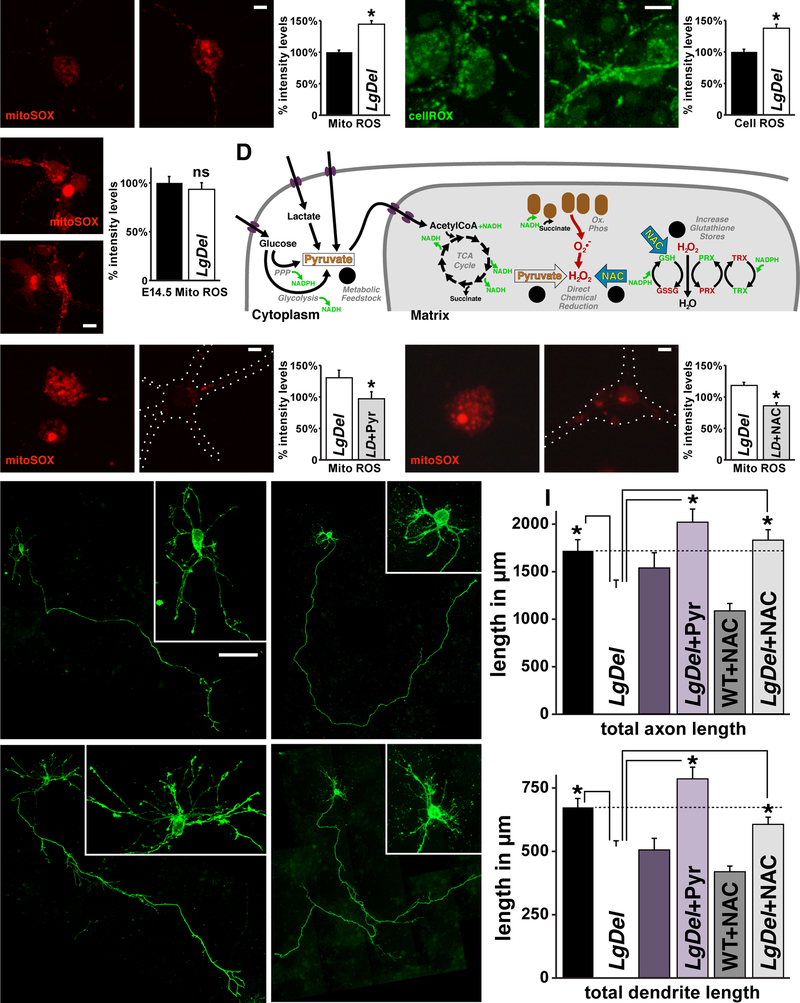
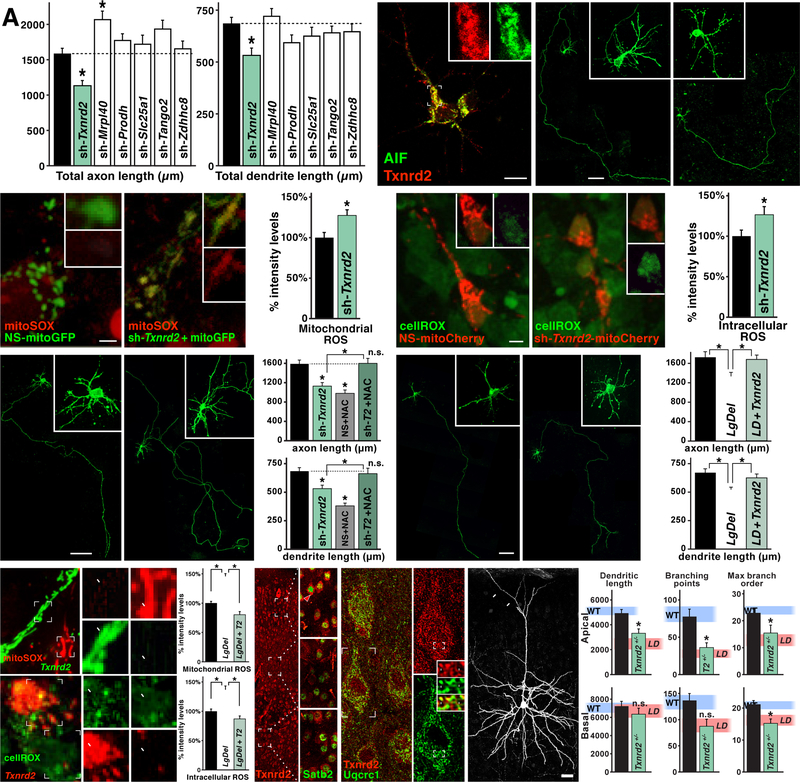
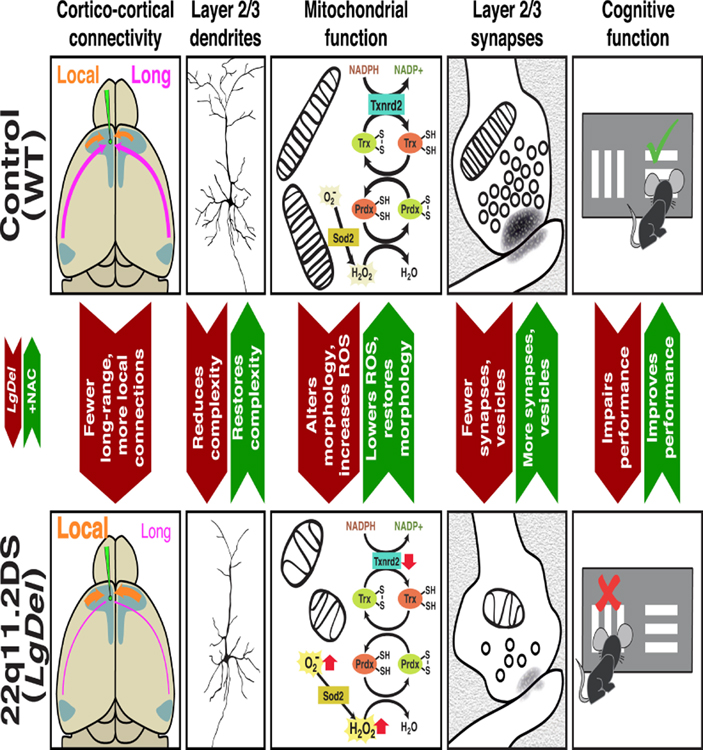
Under-connectivity between cerebral cortical association areas may underlie cognitive deficits in neurodevelopmental disorders, including the 22q11.2 deletion syndrome (22q11DS). Using the LgDel 22q11DS mouse model, we assessed cellular, molecular, and developmental origins of under-connectivity and its consequences for cognitive function. Diminished 22q11 gene dosage reduces long-distance projections, limits axon and dendrite growth, and disrupts mitochondrial and synaptic integrity in layer 2/3 but not 5/6 projection neurons (PNs). Diminished dosage of Txnrd2, a 22q11 gene essential for reactive oxygen species catabolism in brain mitochondria, recapitulates these deficits in WT layer 2/3 PNs; Txnrd2 re-expression in LgDel layer 2/3 PNs rescues them. Anti-oxidants reverse LgDel- or Txnrd2-related layer 2/3 mitochondrial, circuit, and cognitive deficits. Accordingly, Txnrd2-mediated oxidative stress reduces layer 2/3 connectivity and impairs cognition in the context of 22q11 deletion. Anti-oxidant restoration of mitochondrial integrity, cortical connectivity, and cognitive behavior defines oxidative stress as a therapeutic target in neurodevelopmental disorders.
Keywords: autism spectrum disorders; cortical projection neurons; mitochondria; neurodevelopmental disorders; reactive oxygen species; schizophrenia; under-connectivity.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures




Comment in
-
Mitochondrial Function in 22q11 Deletion Syndrome.Neuron. 2019 Jun 19;102(6):1089-1091. doi: 10.1016/j.neuron.2019.05.052. Neuron. 2019. PMID: 31220439
References
-
- Ahlemeyer B, and Baumgart-Vogt E (2005). Optimized protocols for the simultaneous preparation of primary neuronal cultures of the neocortex, hippocampus and cerebellum from individual newborn (P0.5) C57Bl/6J mice. J Neurosci Methods 149, 110–120. - PubMed
-
- Alcamo EA, Chirivella L, Dautzenberg M, Dobreva G, Farinas I, Grosschedl R, and McConnell SK (2008). Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 57, 364–377. - PubMed
-
- Aruoma OI, Halliwell B, Hoey BM, and Butler J (1989). The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free radical biology & medicine 6, 593–597. - PubMed
-
- Bannister AP (2005). Inter- and intra-laminar connections of pyramidal cells in the neocortex. Neuroscience research 53, 95–103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
